6.1. La matriz extracelular: concepto y composición
La matriz extracelular (MEC) es el conjunto de macromoléculas que las células del tejido conjuntivo secretan al espacio intercelular. Es una estructura dinámica que:
- Regula el tráfico de nutrientes y señales entre células.
- Determina las propiedades mecánicas del tejido.
- Actúa como reservorio de factores de crecimiento.
La MEC del tejido conjuntivo tiene dos componentes principales: las fibras y la sustancia fundamental.
- Las fibras son proteínas polimerizadas que forman estructuras filamentosas con propiedades mecánicas definidas.
- La sustancia fundamental es una fase acuosa de gel viscoso que ocupa el espacio entre las fibras.
Existen tres tipos de fibras en el tejido conjuntivo: 1) las fibras de colágeno, 2) las fibras de reticulina y 3) las fibras elásticas. Cada una tiene composición, morfología y función propias.
6.2. Fibras de colágeno
El colágeno es la proteína más abundante del organismo humano, con una representación del 25–30% de la masa proteica total. Sus fibras son el componente más abundante de la MEC del tejido conjuntivo y su función principal es resistir las fuerzas de tracción.
6.2.1. Morfología y organización jerárquica
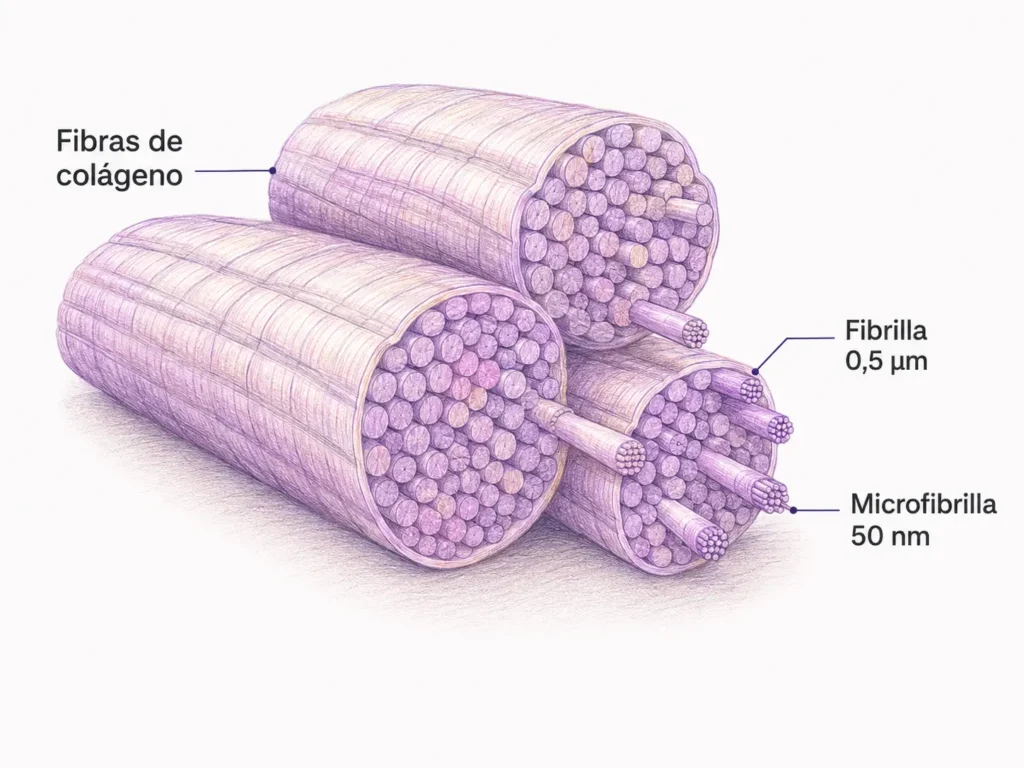
Las fibras de colágeno tienen una organización jerárquica que va desde la molécula hasta el haz macroscópico:
- La unidad molecular es el tropocolágeno:
- Tres cadenas polipeptídicas (cadenas α) enrolladas en una triple hélice dextrógira.
- La secuencia repetitiva Gly-X-Y caracteriza a todas las cadenas α, donde X suele ser prolina e Y hidroxiprolina.
- La glicina en cada tercer residuo es imprescindible porque es el único aminoácido suficientemente pequeño para ocupar el interior de la triple hélice.
- Las moléculas de tropocolágeno miden 280–300 nm de longitud y son polarizadas (tienen un extremo amino-terminal y un extremo carboxilo-terminal).
- Las moléculas de tropocolágeno se agregan lateralmente formando microfibrillas (~50 nm de diámetro). El rasgo más importante de esta agregación es el cabalgamiento escalonado:
- Cada molécula se desplaza ¼ de su longitud respecto a la adyacente y deja un espacio de ~40 nm entre su extremo y el inicio de la siguiente.
- Este patrón de solapamiento (zona de brecha + zona de solapamiento alternadas) genera la estriación periódica característica del colágeno al ME, con una periodicidad de 64–68 nm.
- Las microfibrillas se agrupan en fibrillas (0,2–0,5 µm), rodeadas por una membrana de mucopolisacáridos (membrana de Henle).
- Las fibrillas forman fibras (1–10 µm), visibles al MO.
- Las fibras se agrupan en haces, visibles macroscópicamente como cordones blanco-anacarados.
Al microscopio óptico las fibras de colágeno son:
- Eosinófilas con HE, pero dado que el citoplasma de los fibroblastos también es eosinófilo, las fibras no se distinguen bien de las células.
- Para visualizarlas se usan tinciones específicas:
- Tricrómico de Masson (colágeno en azul).
- Tricrómico de Mallory (verde).
- Van Gieson (rojo).
- Con microscopio de luz polarizada se aprecia birrefringencia longitudinal por la organización cristalina de las fibrillas.
Al microscopio electrónico las fibrillas son perfectamente visibles con su estriación periódica de 64–68 nm, alternando bandas claras y oscuras.
6.2.2. Génesis del colágeno
La síntesis del colágeno ocurre en dos fases secuenciales e inseparables.
- Fase intracelular: el fibroblasto sintetiza las cadenas α en los ribosomas del RER.
- En el interior del RER se producen modificaciones postraduccionales críticas: hidroxilación de prolina e hidroxilación de lisina (dependientes de vitamina C como cofactor), y glicosilación.
- Las tres cadenas se ensamblan en triple hélice y se empaquetan en el Golgi, donde se secretan como procolágeno: la molécula de tropocolágeno con telopéptidos de registro en ambos extremos que impiden la polimerización prematura.
- Fase extracelular (fibrilogénesis): peptidasas específicas del espacio extracelular (procolágeno peptidasa N y C) escindan los telopéptidos del procolágeno.
- En cuanto se liberan los telopéptidos, el tropocolágeno se polimeriza espontáneamente en microfibrillas, fibrillas y fibras.
- Esta polimerización obligatoriamente extracelular es un mecanismo de seguridad. Si el colágeno polimerizara dentro de la célula, destruiría el citoplasma.
La síntesis de las cadenas α, la hidroxilación dependiente de vitamina C y la regulación génica de los distintos tipos de colágeno se desarrollan en Bioquímica.
El escorbuto es la deficiencia de vitamina C. Sin vitamina C, la prolina e hidroxiprolina no se hidroxilan correctamente y el colágeno sintetizado es inestable y no se polimeriza. El resultado es fragilidad capilar, mala cicatrización, sangrado gingival y pérdida de dientes: manifestaciones directas del fallo en la síntesis de colágeno.
6.2.3. Tipos de colágeno
Se han descrito más de 30 tipos de colágeno según la combinación de cadenas α. Los más relevantes para Histología General son:
- Colágeno tipo I: el más abundante del organismo. Forma fibras de 1–10 µm organizadas en haces. Se localiza en dermis, hueso, tendones y dentina. Composición: [α₁(I)]₂α₂.
- Colágeno tipo II: forma fibrillas de 0,5 µm dispuestas en red. Tiene el mismo índice de refracción que la matriz cartilaginosa, por lo que no se distingue al ME en cortes de cartílago. Se localiza en cartílago hialino, núcleo pulposo de los discos intervertebrales y humor vítreo. Composición: [α₁(II)]₃.
- Colágeno tipo III: forma las fibras de reticulina (ver apartado 6.3). Se localiza en órganos hematopoyéticos, hígado, pulmón y paredes vasculares.
- Colágenos que forman mallas (tipos IV, VII, X): no se polimerizan en fibras sino en redes entretejidas. Son los colágenos de la membrana basal, desarrollados en el Tema 7 · Membrana basal y variedades del tejido conjuntivo
| Tipo | Estructura | Localización principal |
|---|---|---|
| I | Fibras (1–10 µm) | Dermis, hueso, tendón, dentina |
| II | Fibrillas en red (0,5 µm) | Cartílago hialino, núcleo pulposo, humor vítreo |
| III | Fibras reticulares finas | Órganos hematopoyéticos, hígado, vasos |
| IV | Malla entretejida | Membrana basal (lámina densa) |
| VII | Fibras de anclaje | Membrana basal (lámina reticular) |
6.3. Fibras de reticulina
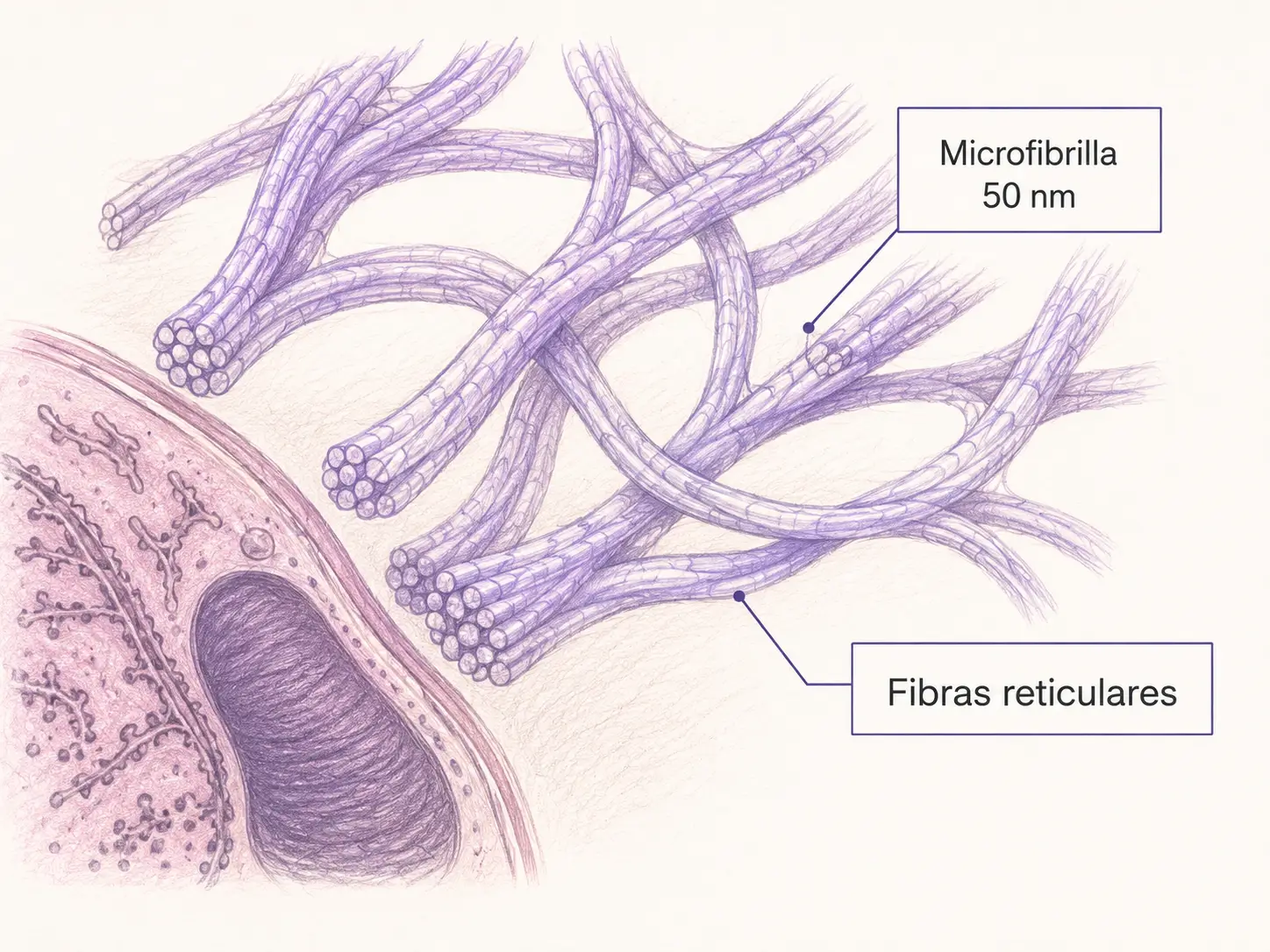
Las fibras de reticulina están formadas por colágeno tipo III con un recubrimiento de proteoglicanos y glicoproteínas que les confiere propiedades tintoriales especiales. Son una variante morfológica del colágeno.
- Se organizan en fibrillas muy finas (≤ 0,5 µm) que forman mallas anastomóticas de sostén.
- Esta organización en red tridimensional las hace ideales para sostener estructuras que necesitan flexibilidad sin rigidez.
- Rodean las células musculares (endomisio), las fibras nerviosas (endoneuro), los adipocitos y forman el estroma de órganos glandulares (hígado) y hematolinfopoyéticos (bazo, ganglios linfáticos, médula ósea).
Al MO no se tiñen con HE. Se identifican por dos propiedades específicas:
- Son PAS positivas (por el recubrimiento de glucoproteínas).
- Sobre todo son argirófilas: se tiñen en negro con la impregnación argéntica de Gomori, que las hace perfectamente visibles como finas mallas negras sobre fondo claro.
Al ME son indistinguibles del colágeno tipo I, con la misma estriación periódica de 64–68 nm.
6.4. Fibras elásticas
Las fibras elásticas confieren elasticidad al tejido conjuntivo. Pueden deformarse bajo tensión y recuperar su forma original al cesar la fuerza. Son resistentes a la ebullición pero muy sensibles a las elastasas.
6.4.1. Morfología y composición de las fibras elásticas
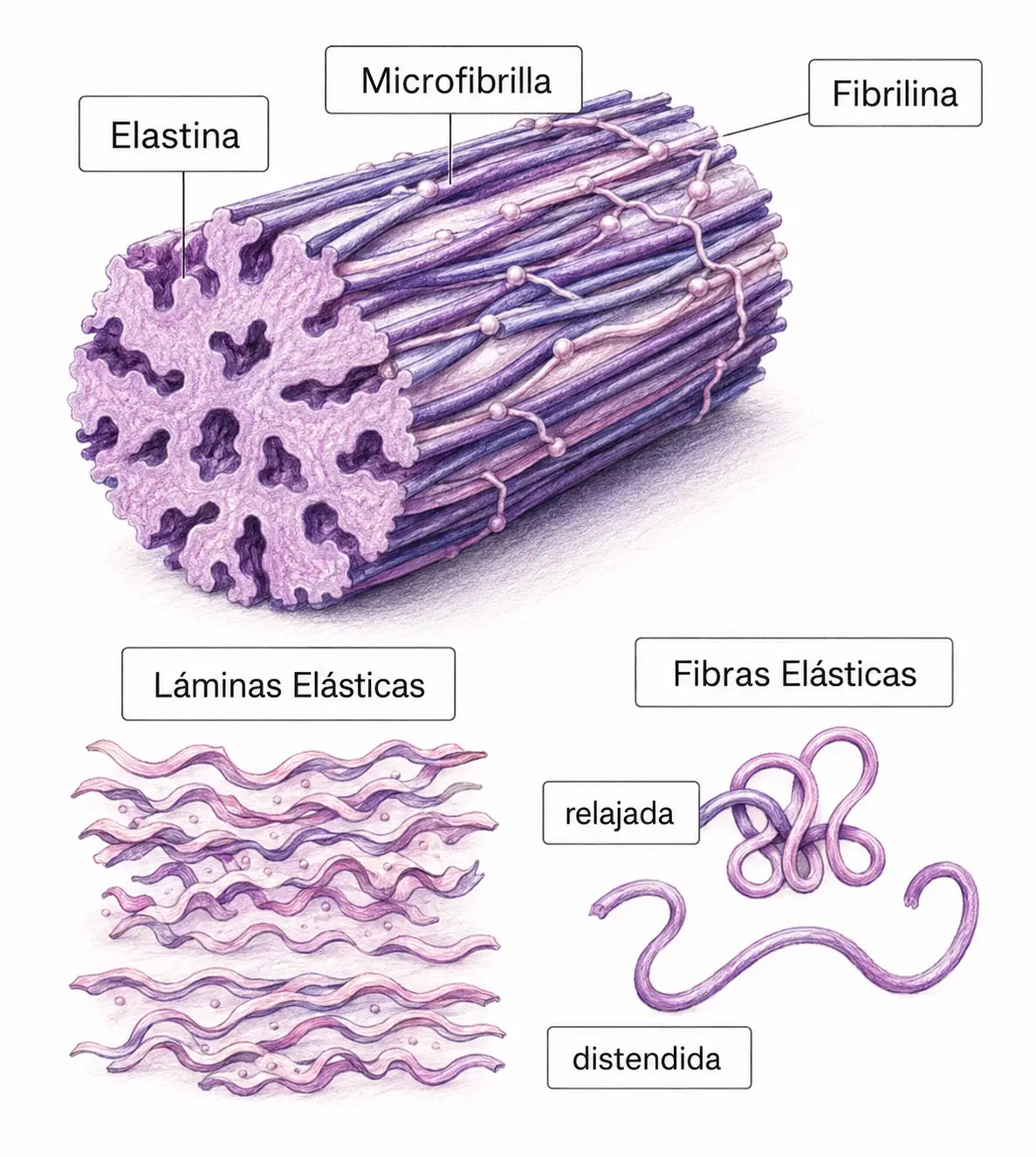
Las fibras elásticas tienen una organización centro-periférica de dos componentes:
- El núcleo central está formado por elastina: una proteína con aminoácidos hidrofóbicos (glicina, valina, alanina) y dos aminoácidos exclusivos, desmosina e isodesmosina, que forman entrecruzamientos covalentes entre moléculas y son la base molecular de la elasticidad. La elastina no tiene la secuencia Gly-X-Y del colágeno ni forma triple hélice.
- La periferia está formada por microfibrillas de fibrilina (10 nm de diámetro), cadenas polipeptídicas de secuencias no repetitivas que forman un andamiaje periférico.
Al MO con HE las fibras elásticas son invisibles o apenas perceptibles. Para su identificación se usan tinciones específicas:
- Para su identificación se usan tinciones específicas: orceína (rojo-pardo) y fucsinas (rojo).
- Son fibras más finas que las de colágeno (0,2 µm de diámetro) y se disponen en mallas bifurcadas.
- Con microscopio de luz polarizada no muestran birrefringencia (a diferencia del colágeno).
Al ME se aprecia claramente la organización interna: microfibrillas de fibrilina en la periferia y material amorfo de elastina en el centro.
6.4.2. Génesis y tipos de fibras elásticas
La génesis de las fibras elásticas sigue una secuencia que determina directamente su organización espacial.
La fibrilina se sintetiza y secreta primero y se sitúa en la periferia; la elastina se sintetiza después y se deposita en el interior, rellenando el andamiaje de fibrilina.
El síndrome de Marfan se produce por mutaciones en el gen de la fibrilina-1. La fibrilina defectuosa altera la integridad de las fibras elásticas en la aorta, el ojo (suspensión del cristalino mediante la zónula de Zinn) y el esqueleto. La consecuencia más grave es la dilatación y disección aórtica.
Existen tres tipos que representan estadios de maduración progresiva:
- Oxitalan: solo contiene microfibrillas de fibrilina, sin elastina. Es la forma más inmadura. Se localiza en la zónula de Zinn (inserción del cristalino) y en el ligamento periodontal.
- Elaunina: contiene fibrilina y una pequeña cantidad de elastina. Se encuentra en la dermis profunda.
- Fibras elásticas maduras: contienen fibrilina periférica y núcleo central abundante de elastina. Según su organización en el tejido se distinguen:
- En plexos (pulmón, dermis superficial).
- En haces fasciculares (ligamento amarillo intervertebral).
- En láminas concéntricas (pared aórtica y grandes arterias).
| Tipo | Composición | Localización |
|---|---|---|
| Oxitalan | Solo fibrilina | Zónula de Zinn, ligamento periodontal |
| Elaunina | Fibrilina + poca elastina | Dermis profunda |
| Maduras (plexos) | Fibrilina + elastina | Pulmón, dermis superficial |
| Maduras (fascicular) | Fibrilina + elastina | Ligamento amarillo intervertebral |
| Maduras (láminas) | Fibrilina + elastina | Aorta, grandes arterias |
6.5. Sustancia fundamental
La sustancia fundamental es la fase no fibrilar de la MEC: Consisten en un gel acuoso viscoso que ocupa el espacio entre las fibras y entre las células. Es el medio a través del cual difunden nutrientes, gases, iones y señales moleculares entre los capilares y las células del tejido.
Al MO convencional la sustancia fundamental es prácticamente invisible:
- El agua que la hidrata se pierde durante la fijación y la inclusión en parafina, y lo que queda son zonas pálidas y homogéneas.
- Para visualizarla se requiere congelación del tejido y tinciones específicas: PAS (positiva por los carbohidratos de los proteoglicanos) y azul alcián (positiva por los proteoglicanos ácidos).
- Las técnicas de inmunofluorescencia son las más precisas.
Al ME se aprecian densificaciones amorfas de 10–20 nm sin estructura definida.
Está compuesta por agua, sales minerales, proteoglicanos y glicoproteínas de adhesión.
6.5.1. Proteoglicanos
Los proteoglicanos están formados por una proteína central (core protein) a la que se unen covalentemente cadenas de glicosaminoglicanos (GAG, antes llamados mucopolisacáridos). Los GAG son cadenas lineales repetitivas de disacáridos, frecuentemente sulfatadas, altamente electronegativas, que atraen cationes (Na⁺) y retienen grandes cantidades de agua. Esta hidrofilia determina que la sustancia fundamental actúe como un gel que regula el tamaño de los poros de la matriz y el tráfico molecular.
Los principales glicosaminoglicanos son:
- Ácido hialurónico: es el único GAG que existe libre, sin unirse a proteína core (técnicamente no es un proteoglicano). Es el de mayor peso molecular y mayor capacidad de retención de agua.
- Forma la cadena central de los grandes agregados de proteoglicanos.
- Muy abundante en líquido sinovial, humor vítreo y cordón umbilical.
- Algunas bacterias producen hialuronidasa, que destruye el ácido hialurónico y facilita la invasión tisular.
- Condroitín sulfato (4 y 6): muy abundante en la matriz cartilaginosa. Forma parte del agrecán junto con el queratán sulfato.
- Queratán sulfato: localizado en cartílago y córnea.
- Dermatán sulfato: predominante en la dermis cutánea.
- Heparán sulfato: ampliamente distribuido, especialmente en órganos como hígado y pulmón, y en la membrana basal (perlecán).
Los monómeros de proteoglicano (proteína core + GAGs) se agregan al ácido hialurónico mediante proteínas de enlace para formar grandes agregados de proteoglicanos. El agrecán (condroitín + queratán sulfato) es el prototipo de estos agregados en el cartílago.
6.5.2. Glicoproteínas de adhesión
Las glicoproteínas de adhesión tienen una composición diferente a los proteoglicanos: predomina la proteína sobre el azúcar, las cadenas de polisacáridos son cortas, ramificadas, no repetitivas y no sulfatadas. Su función principal es actuar como puentes moleculares entre las células y los distintos componentes de la matriz.
- Fibronectina: molécula en forma de dímero unido por puentes disulfuro. Tiene dominios de unión específicos para colágeno I y III, para laminina, para integrinas de la membrana celular y para fibrina. Es la principal molécula de adhesión del tejido conjuntivo y juega un papel crucial en la migración celular durante la reparación de heridas.
- Laminina: morfología cruciforme. Es el componente principal de la lámina rara externa de la membrana basal. Tiene dominios de unión para integrinas, fibronectina, colágeno IV y entactina-nidogen. Es la molécula de adhesión específica del epitelio con su membrana basal.
- Entactina-nidogen: localizada en la membrana basal, forma puentes entre laminina y colágeno IV.
- Osteonectina: específica del hueso, donde participa en la mineralización de la matriz ósea.
- Condronectina: específica del cartílago, media la adhesión de los condrocitos a la matriz.
La membrana basal, sus capas (lámina rara externa, lámina densa, lámina reticular) y su composición molecular completa se desarrollan en T7 — Membrana basal y variedades del tejido conjuntivo.
