8.1. Concepto y funciones del tejido adiposo
El tejido adiposo es una variedad de tejido conjuntivo especializado en el almacenamiento de lípidos. Su célula característica es el adipocito, derivado de la célula mesenquimática indiferenciada. Representa entre el 15 y el 20% del peso corporal en un adulto sano, con variaciones importantes según sexo, edad y estado nutricional.
Durante décadas se consideró un tejido de relleno metabólicamente pasivo. Esta visión es incorrecta. El tejido adiposo es un órgano endocrino activo con funciones múltiples:
- Reserva energética. Los triglicéridos almacenados en los adipocitos constituyen la mayor reserva energética del organismo, con una densidad calórica muy superior a la del glucógeno.
- Homeotermia. El tejido adiposo participa activamente en el intercambio energético y en el mantenimiento de la temperatura corporal, especialmente la grasa parda mediante termogénesis.
- Protección mecánica. La grasa periorbitaria, palmar, plantar y periarticular actúa como amortiguador mecánico en zonas de presión o fricción.
- Función endocrina. Los adipocitos sintetizan y secretan adipocinas: leptina (señal de saciedad y reguladora del balance energético), adiponectina (sensibilizadora a la insulina y antiinflamatoria), resistina y otras. La disfunción de esta secreción es central en la fisiopatología de la obesidad y el síndrome metabólico.
- Respuesta a señales neuroendocrinas. El tejido adiposo expresa receptores para ACTH, insulina, adrenalina y noradrenalina, que regulan la lipólisis y la lipogénesis.
La regulación hormonal del metabolismo lipídico, incluyendo el mecanismo de acción de la leptina y la adiponectina sobre el hipotálamo y el músculo, se desarrolla en Fisiología General y en Bioquímica.
Se distinguen dos tipos de tejido adiposo con morfología, distribución y función claramente diferenciadas.
8.2. Grasa unilocular (blanca o amarilla)
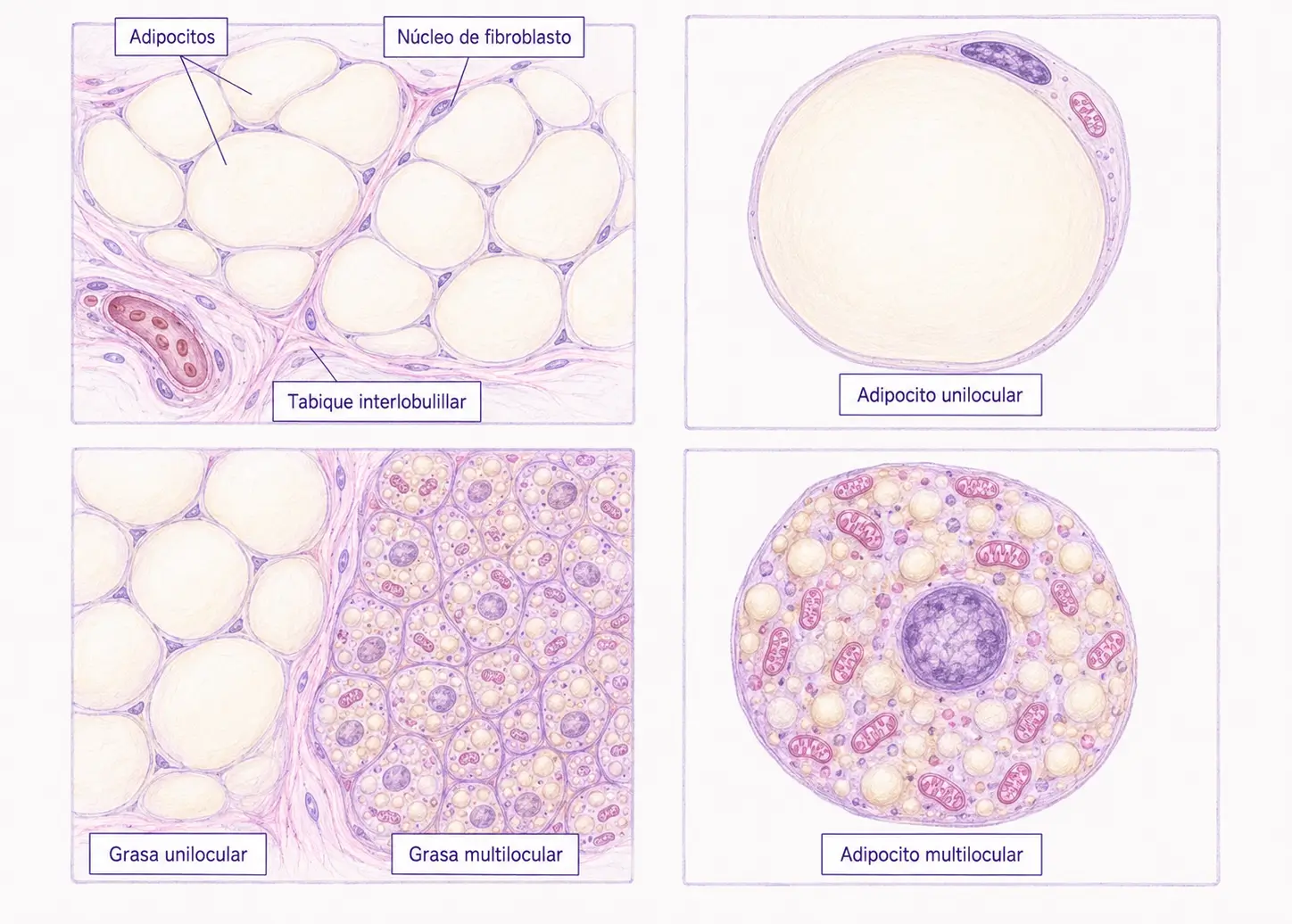
La grasa unilocular es el tipo predominante en el adulto. Debe su nombre a que cada adipocito contiene una única vacuola lipídica que ocupa casi todo el volumen celular. También se denomina grasa blanca o amarilla, por su color macroscópico en fresco, que varía de blanco a amarillento según el contenido de carotenoides de la dieta.
8.2.1. Organización macroscópica
En fresco, el tejido adiposo unilocular presenta una organización lóbulo-lobulillar: una red de fibras de reticulina (colágeno III) forma tabiques que dividen el tejido en lóbulos y lobulillos. Esta estructura alberga los abundantes vasos sanguíneos y fibras nerviosas que necesita el tejido para sus funciones metabólicas.
8.2.2. Morfología al microscopio óptico
La imagen del tejido adiposo blanco al MO varía radicalmente según la técnica de procesado:
- Procesado en parafina (imagen negativa): el lípido se disuelve durante la deshidratación con alcoholes y aclarado con xilol. El resultado es una imagen «negativa»:
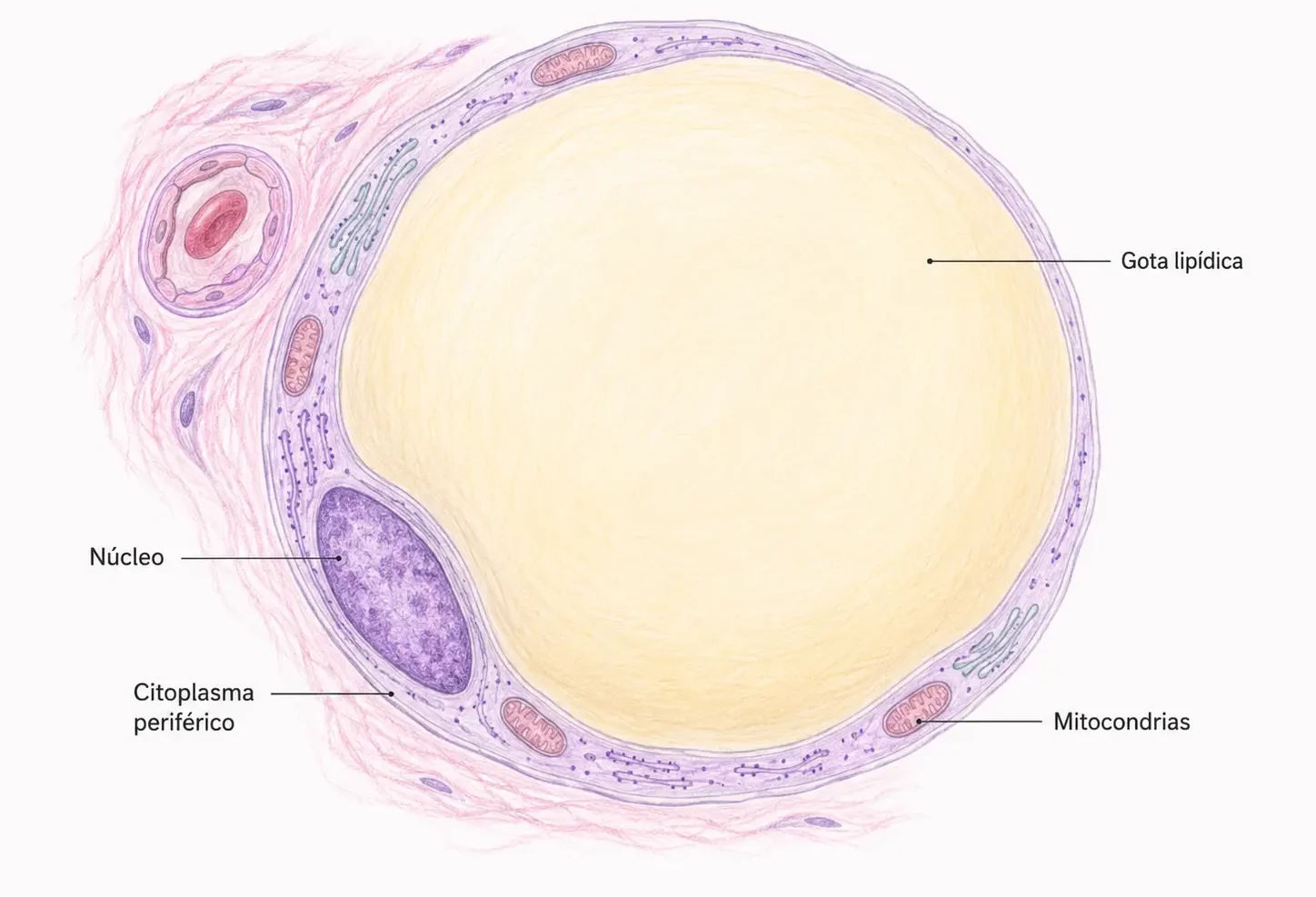
- Las células aparecen pálidas, con una gran vacuola central vacía y un núcleo desplazado a la periferia, aplastado y de cromatina condensada.
- El citoplasma queda reducido a un delgado anillo eosinófilo.
- Esta imagen en anillo de sello es el aspecto habitual del adipocito en los cortes histológicos de rutina. Cada célula está rodeada de una lámina externa similar a la membrana basal.
- Procesado por congelación (imagen positiva): el tejido se congela directamente antes del procesado, preservando el contenido lipídico. Con colorantes liposolubles —negro sudán, rojo congo o azul de toluidina— la vacuola se tiñe intensamente, mostrando la imagen real de la célula llena de lípido.
Cuando los adipocitos se agrupan, sus paredes se aplanan mutuamente dando una morfología facetada, similar a un mosaico de poliedros.
8.2.3. Morfología al microscopio electrónico
El núcleo es alargado y aplastado, desplazado a la periferia por la vacuola. En los polos del núcleo («conos») se concentran un pequeño aparato de Golgi y las mitocondrias, que son filiformes (alargadas y finas) y escasas.
El citoplasma periférico contiene pocos orgánulos (escasos ribosomas y mitocondrias). La vacuola lipídica ocupa el 90–95% del volumen celular y no está rodeada de membrana propia, la delimitan filamentos intermedios de vimentina de 10 nm dispuestos perpendicularmente a su superficie.
En estados de metabolismo lipídico activo pueden aparecer pequeñas vacuolas secundarias emergiendo de la principal. La membrana plasmática presenta vesículas de pinocitosis y exocitosis. Por fuera de la célula se dispone la lámina externa.
8.2.4. Distribución
La distribución de la grasa blanca varía con la edad y el sexo.
En el niño se desarrolla un panículo adiposo subcutáneo continuo que no existe en el adulto.
En el adulto, la distribución es sexualmente dimórfica. En el hombre predomina en nuca, zona lumbar y deltoides; en la mujer, en mamas, nalgas y cara lateral del muslo.
Hay localizaciones comunes a ambos sexos: mesenterio, epiplón y retroperitoneo. Y hay depósitos resistentes a la pérdida, que persisten incluso en estados de caquexia extrema: la grasa periorbitaria, palmar, plantar y periarticular, cuya función mecánica es esencial para la integridad de esas estructuras.
Un factor regulador del almacenamiento lipídico en la grasa blanca es la lipoproteín lipasa (LPL). La LPL se ancla en el endotelio de los capilares del tejido adiposo y que hidroliza los triglicéridos de las lipoproteínas circulantes (VLDL, quilomicrones), liberando ácidos grasos libres que entran al adipocito para su almacenamiento.
La actividad de la LPL tiene regulación hormonal:
- La insulina la activa (favorece el depósito en el estado postprandial).
- La adrenalina/noradrenalina la inhiben (favorecen la lipólisis).
Este mecanismo regula la capacidad de almacenamiento del tejido adiposo blanco de forma dinámica y regional.
La regulación hormonal de la lipoproteín lipasa, el mecanismo de captación de ácidos grasos por el adipocito y el ciclo completo de lipogénesis y lipólisis se desarrollan en Bioquímica — Metabolismo lipídico.
8.3. Grasa multilocular (parda)
La grasa multilocular o parda es el tipo predominante en el feto y en el recién nacido, donde tiene una función termorreguladora crítica. En el adulto persiste en depósitos residuales específicos.
8.3.1. Morfología al microscopio óptico
En fresco presenta un color pardo-rojizo por dos razones independientes:
- La abundante vascularización: mucho mayor que en la grasa blanca. Aporta el color rojizo de la hemoglobina.
- La densísima población de mitocondrias aporta el tono pardo de sus citocromos.
La inervación simpática y parasimpática también es mucho más rica que en la grasa blanca, aunque no influye en el color.
La organización es lobulillar muy marcada, con un aspecto que recuerda a una glándula.
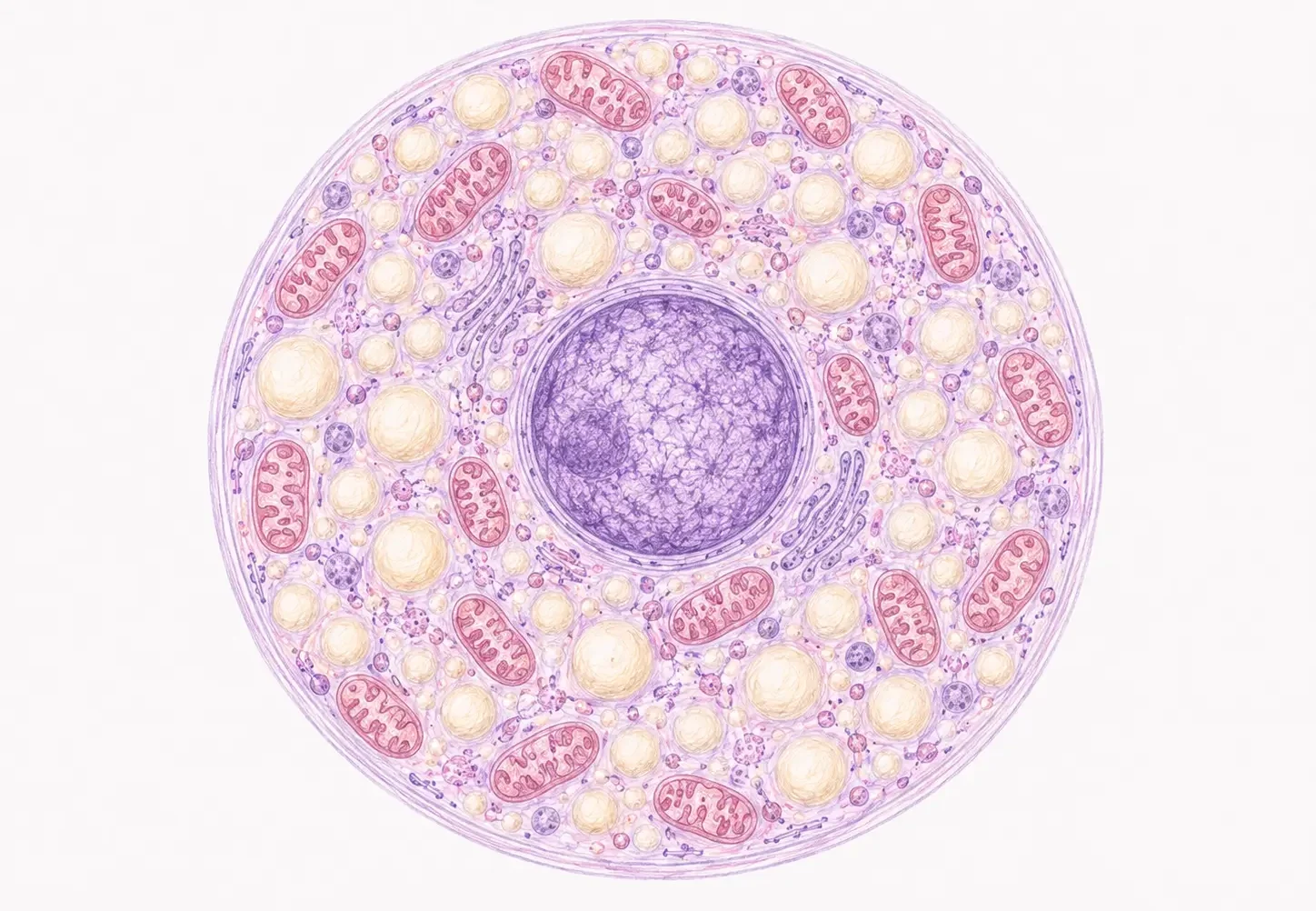
Al MO las células son más pequeñas que los adipocitos blancos (~50 µm).
- El núcleo es redondo, de cromatina laxa, en posición generalmente excéntrica pero no completamente desplazado.
- El citoplasma contiene múltiples vacuolas de distintos tamaños (de ahí el nombre «multilocular»).
- El citoplasma entre las vacuolas es intensamente eosinófilo y granular por las mitocondrias.
8.3.2. Morfología al microscopio electrónico
El núcleo es lateral, de cromatina laxa. Las vacuolas lipídicas son múltiples, sin membrana propia, delimitadas por filamentos de vimentina.
El hallazgo más llamativo es la presencia de centenares de mitocondrias regulares (no filiformes como en la grasa blanca), que son el orgánulo dominante del citoplasma.
Las mitocondrias son la base ultraestructural de la eosinofilia granular visible al MO.
La membrana plasmática presenta también lámina externa y vesículas de pinocitosis/exocitosis.
La clave funcional de la grasa parda es la proteína UCP-1 (termogenina), localizada en la membrana interna mitocondrial. UCP-1 actúa como un canal que disipa el gradiente de protones generado por la cadena respiratoria directamente como calor, sin sintetizar ATP. Este mecanismo permite la termogénesis sin escalofríos, esencial en el neonato y en animales de hibernación.
8.3.3. Distribución
En el recién nacido: hilio renal, axilas, vainas de las carótidas, región pertiroidea e interescapular.
En los primeros días postnatales la grasa multilocular modifica su morfología y adopta el aspecto de la grasa unilocular. Sin embargo, en situaciones de caquexia la grasa de esos depósitos específicos puede retransformarse a su morfología multilocular original, lo que confirma que las dos poblaciones coexisten de forma latente.
Los animales de hibernación tienen una acumulación muy abundante de grasa escapular parda, que cumple una doble función:
- Proporcionar energía durante el ayuno de la hibernación.
- Termorregular el organismo al despertar, evitando los escalofríos que podrían ser peligrosos tras la hipotermia.
Los tumores del tejido adiposo son los más frecuentes de los tumores de partes blandas. Los lipomas y liposarcomas pueden derivar tanto de la grasa blanca como de la parda.
Los tumores de grasa parda se denominan hibernomas. La existencia de tumores diferenciados de cada tipo confirma que las dos variedades proceden de linajes celulares distintos.
8.4. Comparativa grasa blanca vs. grasa parda
| Característica | Grasa unilocular (blanca) | Grasa multilocular (parda) |
|---|---|---|
| Vacuolas | Una grande (90–95% del volumen) | Múltiples, de tamaño variable |
| Núcleo | Periférico, aplastado, condensado | Excéntrico, redondo, cromatina laxa |
| Mitocondrias | Escasas, filiformes | Centenares, regulares (UCP-1) |
| Citoplasma MO | Anillo eosinófilo fino | Abundante, eosinófilo granular |
| Color en fresco | Blanco-amarillento (carotenos) | Pardo-rojizo (citocromos + Hb) |
| Vascularización | Moderada | Muy abundante |
| Inervación | Moderada | Rica (simpática y parasimpática) |
| Función principal | Reserva energética | Termogénesis sin escalofríos |
| Predominio | Adulto | Feto y recién nacido |
| Adipogénesis | Secundaria (perinatal) | Primaria (fetal) |
8.5. Adipogénesis
La adipogénesis es el proceso de diferenciación de las células mesenquimáticas indiferenciadas en adipocitos. Sigue dos vías distintas que originan los dos tipos de grasa.
Formación primaria → grasa multilocular
Se produce durante los primeros meses de la vida fetal. A partir de células mesenquimáticas indiferenciadas surge el lipoblasto, morfológicamente indistinguible de su precursor con técnicas habituales.
Los lipoblastos se agrupan en estructuras pseudoglanduloides que desarrollan en su interior múltiples vacuolas lipídicas pequeñas denominados prolipocitos primarios.
Estos evolucionan a lipocitos primarios con el citoplasma repleto de vacuolas multiloculares. A partir de este punto los pasos son reversibles, lo que explica la retransformación de la grasa blanca a parda en la caquexia.
Formación secundaria → grasa unilocular
Se produce en los últimos días de gestación y el primer mes postnatal. Las células mesenquimáticas y los lipoblastos acumulan vacuolas individuales sin adoptar la disposición pseudoglanduloide. Las vacuolas se fusionan progresivamente hasta originar la gran vacuola única del adipocito unilocular.
El número de adipocitos queda fijado en el primer mes postnatal. Es el único período en que los adipocitos pueden proliferar de forma significativa tras el nacimiento.
A partir de ese momento, la ganancia o pérdida de grasa en el adulto se produce por hipertrofia o atrofia de los adipocitos existentes, no por aumento de su número.
Las obesidades de inicio en la primera infancia por sobrealimentación en ese período crítico generan un número aumentado de adipocitos que persiste de por vida y predispone a la obesidad mórbida. Las obesidades mórbidas sin ese antecedente son generalmente de causa patológica.
