2.1. El tejido epitelial: concepto y propiedades generales
El tejido epitelial es uno de los cuatro tejidos básicos del organismo. Está formado por células íntimamente unidas entre sí que se organizan en láminas o masas compactas. Lo que define a cualquier epitelio, independientemente de su tipo o localización, son cuatro propiedades que siempre se cumplen juntas.
- Las células epiteliales son avasculares. Ningún vaso sanguíneo penetra en el epitelio. Los nutrientes y el oxígeno llegan por difusión desde el tejido conjuntivo subyacente, atravesando la membrana basal. Esta dependencia del conjuntivo limita el grosor máximo que puede alcanzar un epitelio.
- Las células descansan sobre una membrana basal. Esta estructura extracelular, producida conjuntamente por las células epiteliales y el tejido conjuntivo adyacente, actúa como soporte mecánico, barrera selectiva y señal de orientación para la migración y renovación celular. Su composición y función se desarrollan en el T6 — Tejido conjuntivo II.
- Las células epiteliales presentan polaridad morfológica y funcional. Toda célula epitelial tiene un polo apical orientado hacia la luz o la superficie libre, y un polo basal orientado hacia la membrana basal. Esta asimetría no es solo estructural: las proteínas de membrana, los orgánulos y las diferenciaciones de superficie se distribuyen de forma diferente en cada polo, lo que permite que la célula transporte moléculas de forma direccional.
- El espacio intercelular es mínimo o prácticamente inexistente. Las células están unidas entre sí por complejos de unión que sellan el espacio entre ellas, aseguran la cohesión mecánica del tejido y, en muchos casos, forman barreras impermeables.
Tejido epitelial de revestimiento: conjunto de células epiteliales organizadas en láminas continuas, avasculares y polarizadas, que revisten superficies corporales o cavidades internas y se apoyan sobre una membrana basal.
La membrana basal no es simplemente una lámina de sostén: contiene colágeno tipo IV, laminina, fibronectina y proteoglicanos que participan en señalización celular, filtración y regeneración tisular. Su estructura completa se estudia en T6 — Tejido conjuntivo II.
Dentro del tejido epitelial se distinguen dos grandes grupos funcionales: los epitelios de revestimiento, que cubren superficies y cavidades, y los epitelios glandulares, especializados en la síntesis y secreción de sustancias. Este tema aborda los primeros. Los epitelios glandulares se estudian en el Tema 3 · Epitelios glandulares.
2.2. Clasificación de los epitelios de revestimiento
La clasificación de los epitelios de revestimiento se basa en dos criterios morfológicos que se aplican siempre en el mismo orden.
- El primer criterio es el número de capas de células:
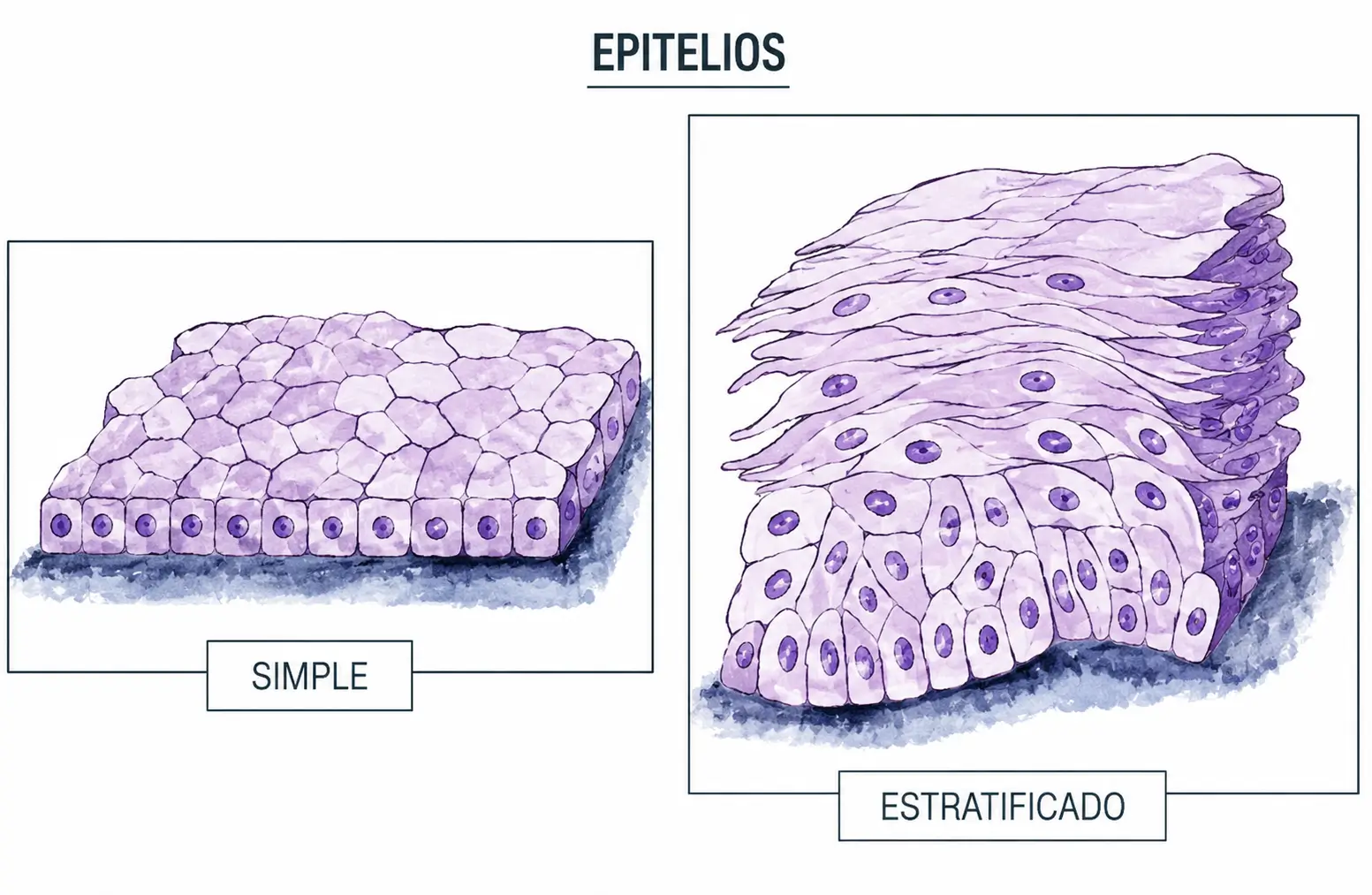
- Epitelio simple (o monoestratificado): si todas las células contactan con la membrana basal y solo existe una capa.
- Epitelio estratificado: si hay dos o más capas. Solo las células del estrato más profundo contactan con la membrana basal, y solo las del estrato más superficial están en contacto con la luz o la superficie.
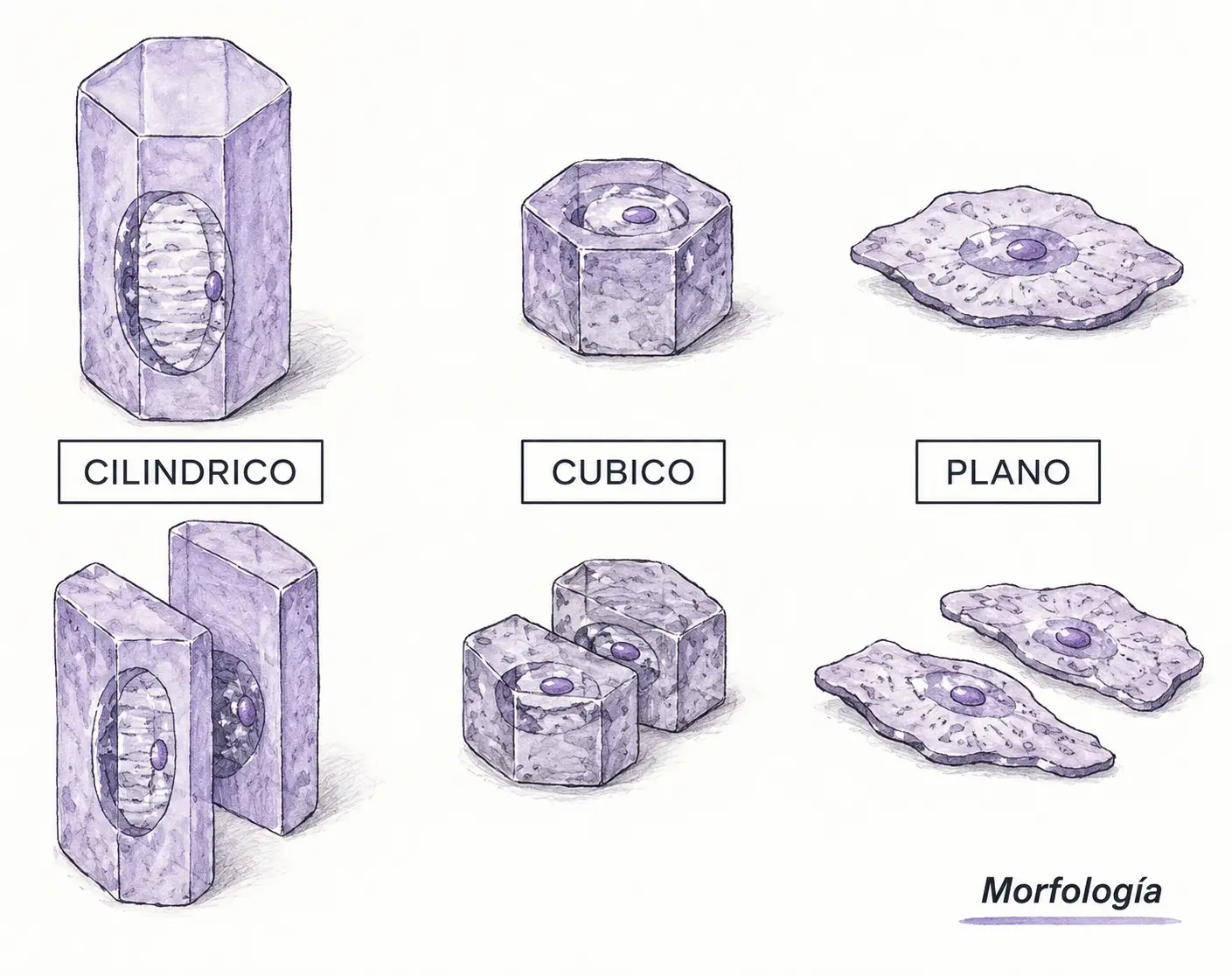
- El segundo criterio es la forma geométrica de las células del estrato más superficial:
- Las células pueden ser planas (más anchas que altas).
- Cúbicas (isodiamátricas).
- Cilíndricas (más altas que anchas, también llamadas prismáticas).
Este segundo criterio define el nombre final del epitelio.
La combinación de ambos criterios genera la nomenclatura: «epitelio simple plano», «epitelio estratificado cúbico», etc.
Hay dos variantes que escapan parcialmente a esta lógica y merecen atención especial: el epitelio pseudoestratificado y el epitelio de transición.
2.3. Tipos de epitelios simples
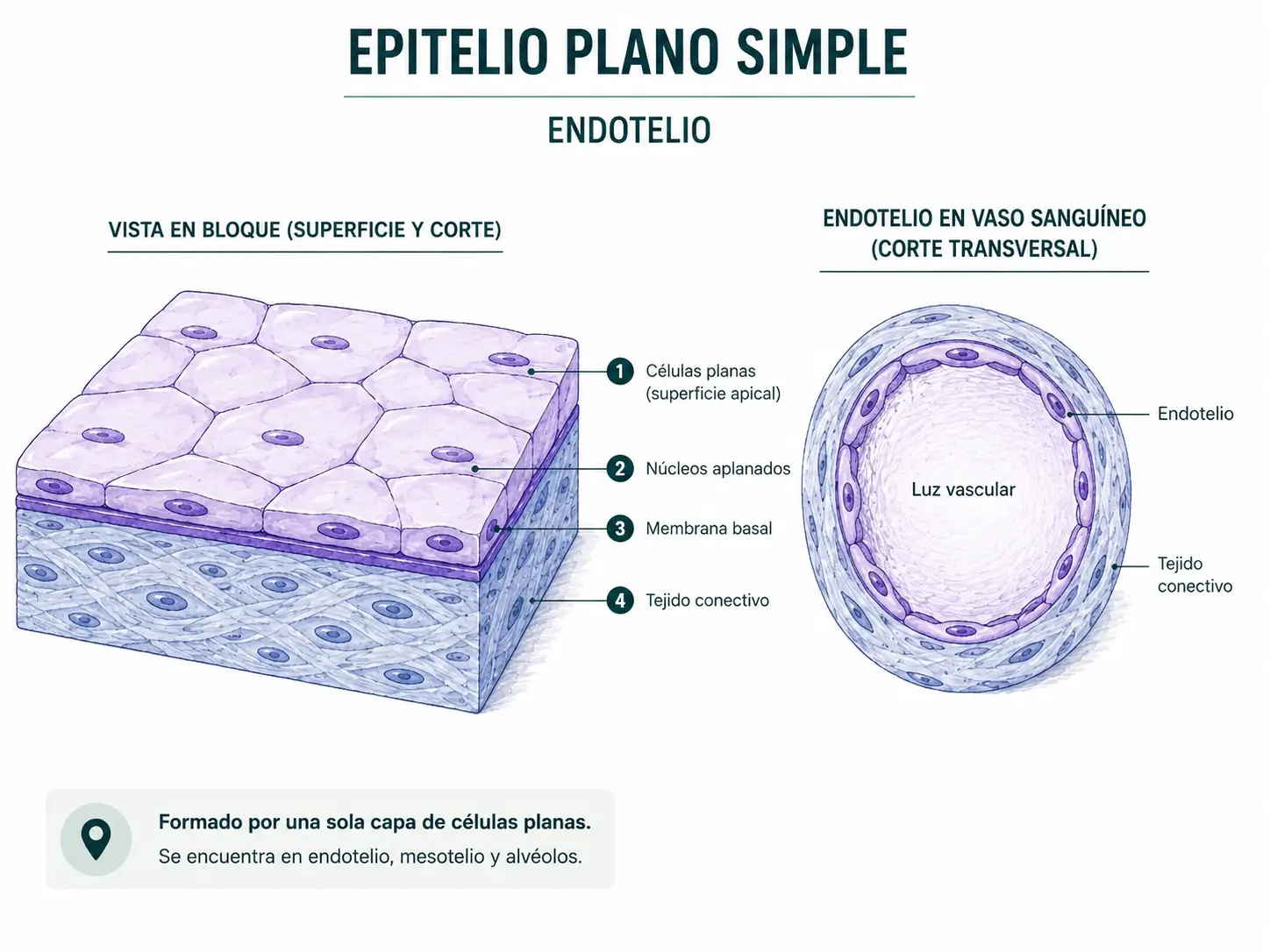
2.3.1. Epitelio simple plano
Está formado por una única capa de células más anchas que altas. Vistas de perfil parecen rectángulos muy aplanados con una pequeña protuberancia central donde se aloja el núcleo. Vistas desde arriba adoptan la forma de polígonos irregulares yuxtapuestos, como un pavimento de adoquines.
Su extrema delgadez favorece el intercambio rápido de gases, iones y nutrientes entre compartimentos. Por eso reviste estructuras donde la difusión o el transporte pasivo son prioritarios:
- Vasos sanguíneos y linfáticos: aquí recibe el nombre de endotelio.
- Cavidades serosas (pleural, pericárdica, peritoneal): recibe el nombre de mesotelio.
- Alvéolos pulmonares: epitelio alveolar.
- Cápsula de Bowman renal.
El endotelio vascular —epitelio simple plano especializado que tapiza todos los vasos— se estudia en profundidad en Histología Especial, dentro del aparato circulatorio.
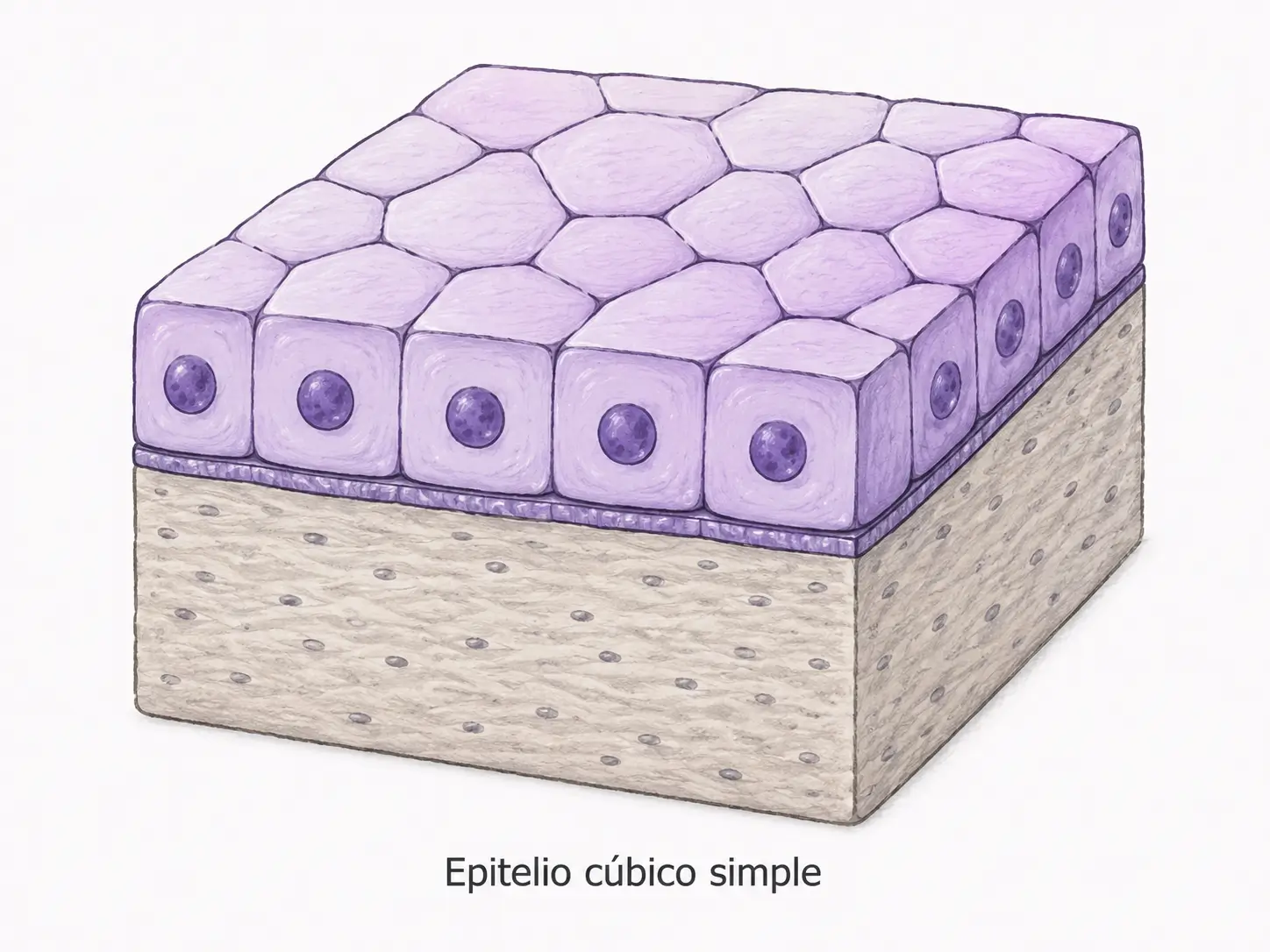
2.3.2. Epitelio simple cúbico
Está formado por una única capa de células isodiamátricas. Su anchura, altura y profundidad son aproximadamente iguales. En corte transversal tienen aspecto de cubo y vistos desde arriba se disponen como hexágonos regulares. El núcleo ocupa una posición central, a la misma altura en todas las células.
Reviste cavidades donde se requiere un tránsito selectivo de moléculas, con una cierta actividad metabólica que justifica la mayor masa citoplasmática respecto al epitelio plano:
- Plexos coroideos (síntesis de líquido cefalorraquídeo).
- Superficie ovárica.
- Conductos de glándulas exocrinas.
- Bronquiolos terminales de pequeño calibre.
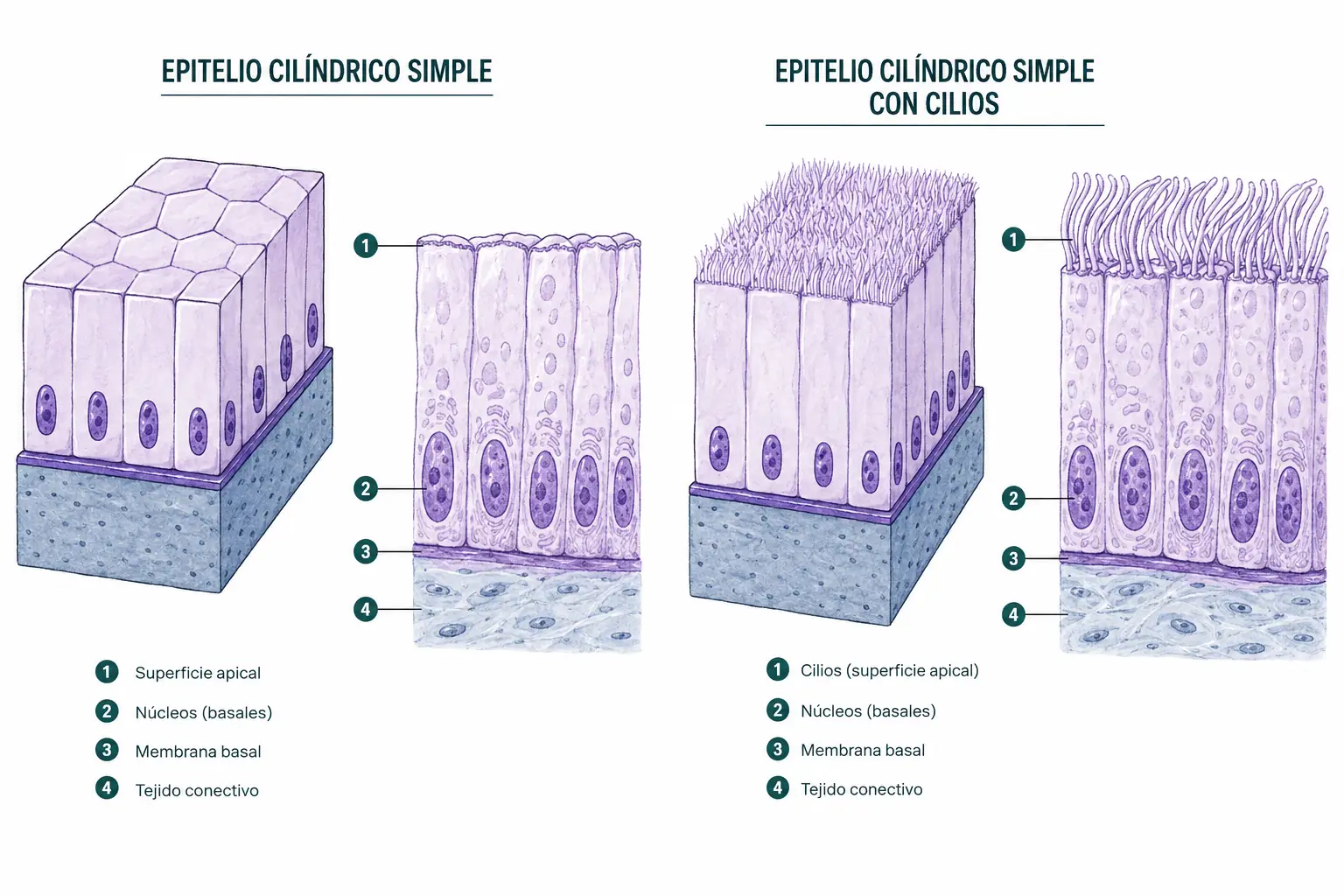
2.3.3. Epitelio simple cilíndrico
Está formado por una capa de células prismáticas, dos a tres veces más altas que anchas. En corte transversal se ven como rectángulos altos. El núcleo ocupa una posición centro-basal, alineado a la misma altura en todas las células.
Es el epitelio más frecuente en superficies con función absortiva o secretora activa. Existe en múltiples variedades según las células especializadas que contiene:
- Variedad intestinal (intestino delgado): alterna células absortivas con microvellosidades apicales y células caliciformes secretoras de moco.
- Variedad gástrica (mucosa del estómago): formada por células secretoras de moco a polo cerrado. No presentan la morfología en cáliz de las células caliciformes: el moco se libera de forma continua y discreta por el polo apical.
- Variedad de la trompa uterina: mezcla de células ciliadas (cinetocilios) que impulsan el ovocito en dirección al útero, y células secretoras de sustrato nutritivo.
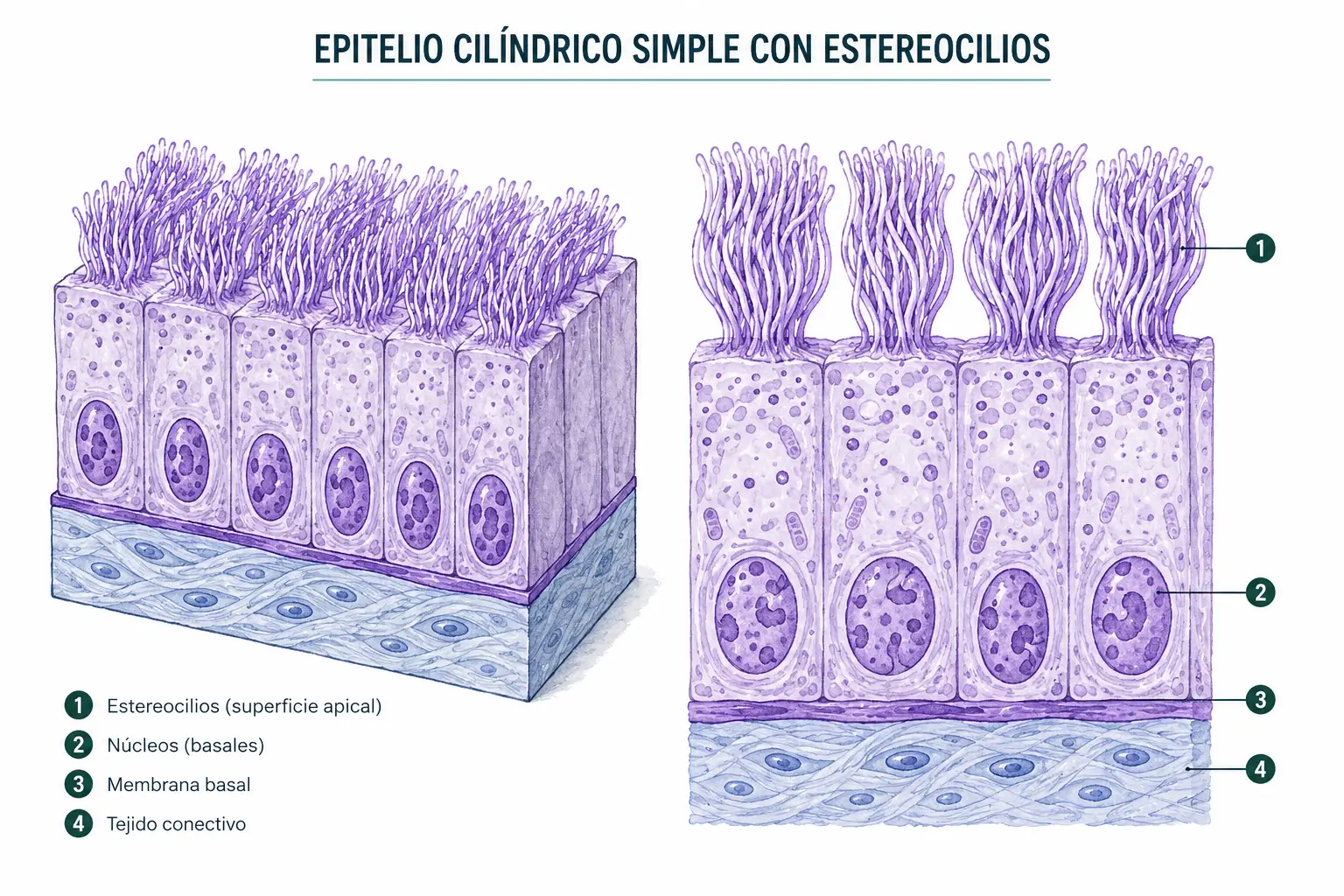
- Variedad del epidídimo: células con estereocilios, que son microvellosidades largas con función absortiva (no cilios móviles, pese al nombre histórico).
2.3.4. Epitelio pseudoestratificado
Es una variedad del epitelio cilíndrico que merece categoría propia por su apariencia característica. Todas sus células contactan con la membrana basal, pero no todas alcanzan la superficie luminal. Dado que los núcleos de células de distinta altura se sitúan a distintos niveles, al microscopio óptico el epitelio parece estratificado. Por eso el prefijo «pseudo», la estratificación es aparente, no real.
En el epitelio pseudoestratificado todas las células tocan la membrana basal. Si alguna célula no la toca, ya no es pseudoestratificado: es un epitelio verdaderamente estratificado. El criterio definitivo es el contacto basal, no la posición de los núcleos.
La variedad más importante es el epitelio respiratorio (tráquea y bronquios principales), que combina células ciliadas, células caliciformes y células basales de reserva.
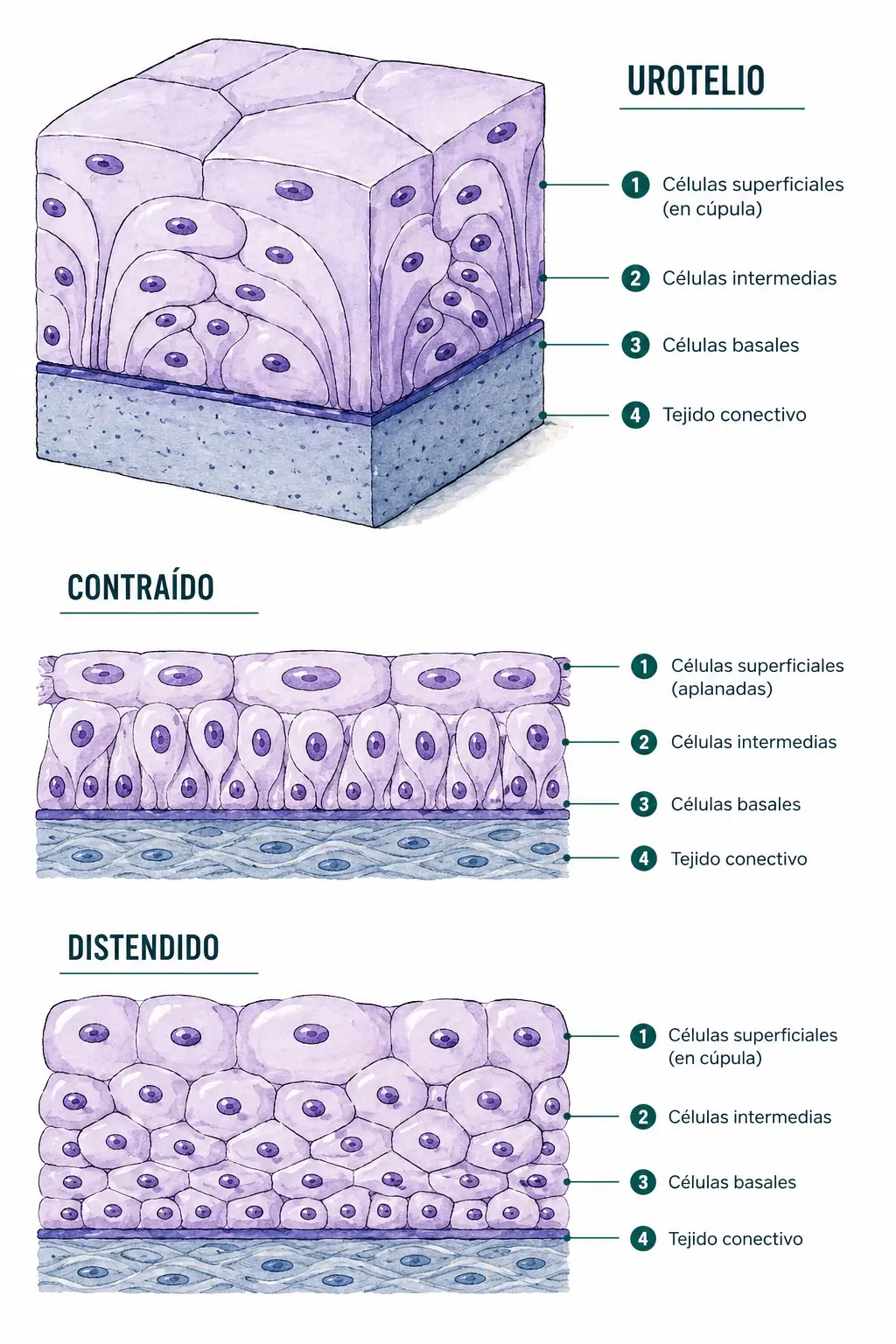
2.3.5. Epitelio de transición (urotelio)
Es una variante especial que tapiza desde los cálices renales hasta la uretra, y cuya característica definitoria es su plasticidad. Cambia de aspecto según el grado de distensión de la pared:
- Cuando la vejiga está vacía o contraída, el epitelio aparece con seis o más capas de células, y las células superficiales son voluminosas, con aspecto redondeado o en cúpula. Se denominan células en paraguas o células de Dögiel. Al microscopio electrónico presentan cisternas apicales en forma de acordeón que se despliegan al distenderse la pared, y una membrana plasmática apical engrosada («placa asimétrica») con una composición lipídica especial que la hace muy impermeable a la orina.
- Cuando la vejiga está distendida y llena, las células en paraguas se aplanan y las capas aparentes se reducen a una o dos.
La organización microscópica completa del urotelio, con sus tres poblaciones celulares y las características ultraestructurales de las células en paraguas, se desarrolla en Histología Especial, dentro del aparato urinario.
El urotelio no cambia realmente el número de células: lo que cambia es su forma y la disposición aparente de las capas. La impermeabilidad a la orina (pH ácido, urea, sales) depende de las placas asimétricas de las células en paraguas, que son la diferenciación funcional más importante de este epitelio.
2.4. Tipos de epitelios estratificados
2.4.1. Epitelio estratificado plano
Es el más frecuente y el más grueso de los epitelios estratificados. Está formado por múltiples capas de células. Las células más basales, en contacto con la membrana basal, son cúbicas o cilíndricas. A medida que ascienden hacia la superficie van aplanándose progresivamente hasta convertirse en células planas en los estratos más superficiales.
El criterio de subclasificación es si acumulan queratina o no.
Epitelio estratificado plano queratinizado (prototipo: epidermis)
Se organiza en cuatro estratos bien diferenciados que representan estadios sucesivos de maduración celular:
- El estrato basal o germinativo es la capa más profunda.
- Está en contacto directo con la membrana basal.
- Se forma por células cúbicas con núcleo redondo y citoplasma basófilo.
- Son las células con mayor capacidad proliferativa del epitelio.
- Contienen gránulos de melanina procedentes de los melanocitos vecinos, localizados justo por encima del núcleo a modo de sombrilla protectora frente a la radiación UV.
- Se anclan a la membrana basal mediante hemidesmosomas y entre sí mediante desmosomas.
- El estrato espinoso (3–5 capas):
- Está formado por células poligonales que a medida que ascienden se van aplanando.
- Su citoplasma es menos basófilo.
- Al microscopio óptico destacan las «espinas» intercelulares, que son los puentes de desmosomas entre células vecinas, visibles porque el fijador retrae el citoplasma y deja en tensión los filamentos de unión.
- Contienen abundantes tonofibrillas (haces de citoqueratina) y, en los estratos superiores, gránulos de Odland o queratinosomas: vesículas rodeadas de membrana que contienen lípidos y serán liberados por exocitosis para formar el cemento intercelular del estrato granuloso.
- El estrato granuloso (2–5 capas):
- Se reconoce al microscopio óptico por la presencia de gránulos de queratohialina, basófilos, sin membrana propia, que son el marcador histológico de este estrato.
- Estos gránulos están rodeados de filagrina, a la que se unen los filamentos de citoqueratina.
- Las células se aplanan progresivamente y los núcleos empiezan a condensarse.
- Los gránulos de Odland se exocitan y los lípidos sellan el espacio intercelular.
- El estrato córneo es el más superficial:
- Está formado por células completamente queratinizadas.
- En el epitelio ortoqueratinizado (epidermis típica), las células del estrato córneo han perdido el núcleo por cariorrexis y se han convertido en escamas planas e inertes, llenas de queratina, que se desprenden continuamente.
- En los epitelios paraqueratinizados (algunas zonas de la mucosa oral) las células mantienen un núcleo picnótico residual.
Epitelio estratificado plano no queratinizado (prototipo: mucosa vaginal, esófago)
Comparte la organización en múltiples capas, pero las células superficiales mantienen siempre el núcleo y no acumulan queratina. Los estratos se denominan de forma diferente:
- El estrato basal es similar al queratinizado. Se compone de células cúbicas basófilas con hemidesmosomas, activas mitóticamente. Sin gránulos de melanina en epitelios de mucosa.
- El estrato parabasal (inmediatamente por encima del basal) muestra células con núcleos redondos y empieza a acumular filamentos de queratina, pero sin formar tonofibrillas. Los gránulos laminados son escasos.
- El estrato intermedio (hasta 5 capas) presenta células aplanadas con núcleo aplanado. Acumulan glucógeno de forma progresiva, que las torna eosinófilas, y reducen los orgánulos de síntesis.
- El estrato superficial contiene células muy aplanadas, muy cargadas de glucógeno, con un núcleo pequeño pero siempre presente.
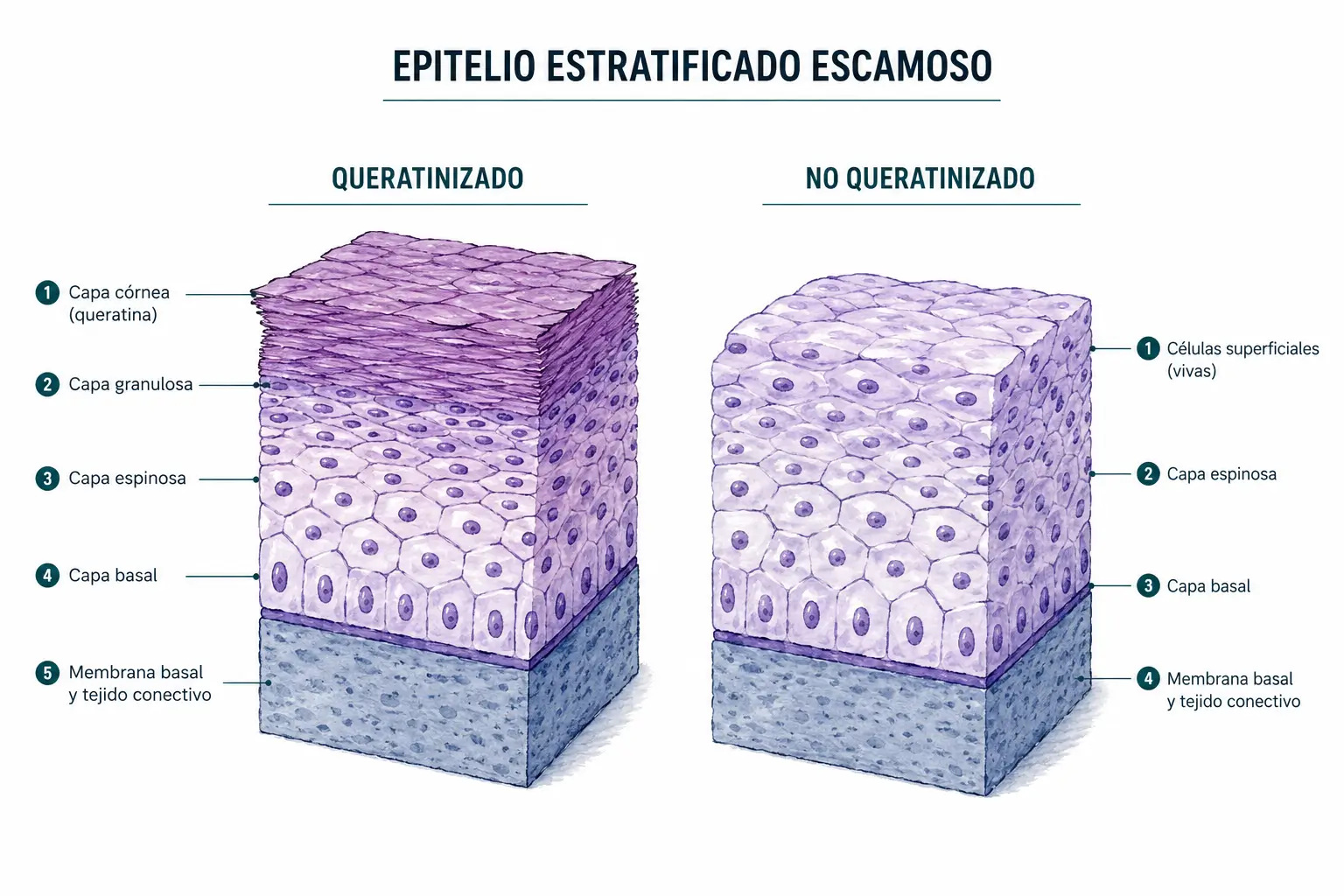
La tabla comparativa de estratos entre el epitelio queratinizado y el no queratinizado es un clásico de examen. La diferencia más importante es la progresión: en el queratinizado, el estrato granuloso es el paso previo a la muerte celular programada y la pérdida del núcleo; en el no queratinizado, ese paso no existe y las células superficiales mantienen viabilidad.
| Estrato | Queratinizado (epidermis) | No queratinizado (esófago, vagina) |
|---|---|---|
| Más superficial | Córneo (sin núcleo, queratina) | Superficial (con núcleo, glucógeno) |
| Intermedio superior | Granuloso (gránulos queratohialina) | Intermedio (glucógeno, achatamiento) |
| Intermedio inferior | Espinoso (desmosomas, tonofibrillas) | Parabasal (queratina sin tonofibrillas) |
| Más profundo | Basal / germinativo (mitosis, melanina) | Basal / germinativo (mitosis) |
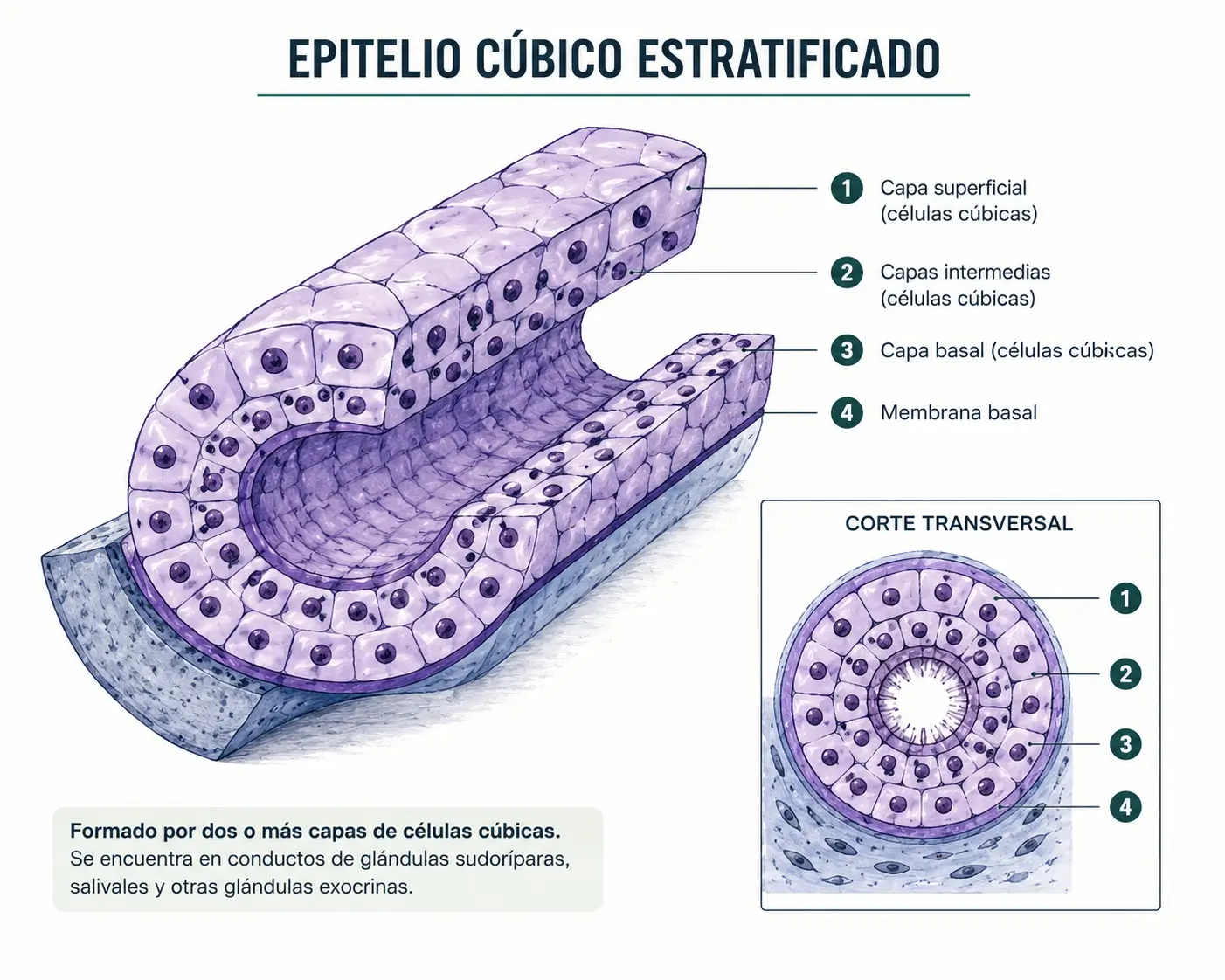
2.4.2. Epitelio estratificado cúbico
Muy poco frecuente en el adulto, más propio del período fetal. Cuando aparece en el adulto se caracteriza por presentar exactamente dos capas de células cúbicas. Se encuentra en la estría vascular del oído interno y en la conjuntiva palpebral.
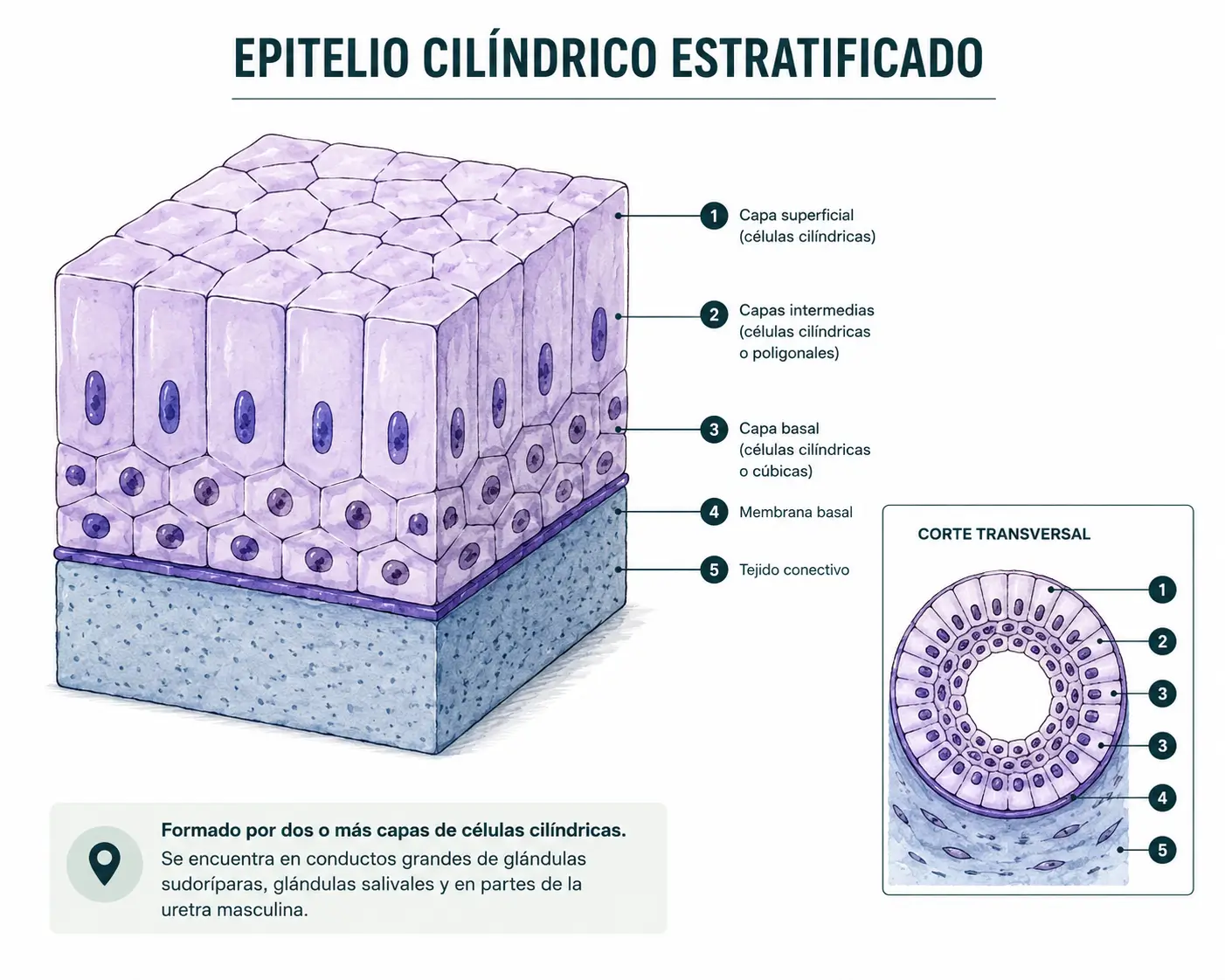
2.4.3. Epitelio estratificado cilíndrico
Igualmente infrecuente. Está formado por dos o tres capas en las que los estratos basales son cúbicos y solo el superficial es cilíndrico. Aparece en la epiglotis, en algunos tramos de conductos glandulares y en la uretra.
2.5. Características celulares de los epitelios de revestimiento
Las células epiteliales comparten una serie de propiedades ultraestructurales que derivan directamente de su función como lámina continua polarizada.
2.5.1. Polaridad y citoesqueleto
Toda célula epitelial tiene un polo basal orientado hacia la membrana basal y un polo apical orientado hacia la luz o la superficie libre.
Esta polaridad se mantiene gracias a un citoesqueleto muy desarrollado, compuesto fundamentalmente por filamentos intermedios de citoqueratina (también llamados tonofilamentos). La citoqueratina es la proteína estructural característica de las células epiteliales y su presencia en un corte histológico o en inmunohistoquímica permite identificar un tejido como epitelial.
2.5.2. Diferenciaciones de la superficie lateral: los complejos de unión
Las células epiteliales desarrollan en su cara lateral estructuras de unión especializadas que sellan el espacio intercelular, aportan cohesión mecánica y permiten la comunicación entre células vecinas. En conjunto se denominan complejo de unión y se organizan de forma ordenada desde el polo apical hacia el basal: uniones ocludentes, uniones adherentes, desmosomas y uniones comunicantes.
Al microscopio óptico con hematoxilina-eosina estas uniones no se individualizan como tales, pero sus efectos son perfectamente visibles:
- Las uniones ocludentes son responsables del sellado que impide el paso paracelular de contenido luminal. Cuando fallan (como en la inflamación intestinal) la barrera epitelial se hace permeable y aparece edema e infiltrado en la lámina propia.
- Las uniones adherentes y los desmosomas son los que mantienen la cohesión del epitelio frente a fuerzas mecánicas. En el epitelio estratificado plano, la pérdida de desmosomas produce la imagen de acantólisis, en la que las células del estrato espinoso se separan entre sí dejando un espacio intercelular visible, como ocurre en el pénfigo vulgar. Los desmosomas también explican las «espinas» intercelulares características del estrato espinoso de la epidermis. Al microscopio óptico, la retracción del citoplasma durante la fijación deja en tensión los puentes desmosómicos entre células vecinas, dando el aspecto espiculado que da nombre al estrato.
- Las uniones comunicantes no tienen expresión morfológica propia al MO, pero su ausencia funcional se manifiesta en pérdida de coordinación entre células epiteliales, relevante en algunos contextos tumorales.
Al microscopio electrónico de transmisión el complejo de unión se identifica con claridad en cortes perpendiculares a la membrana lateral. Los criterios diagnósticos clásicos son:
- La fusión de las hemimembranas externas en las ocludentes.
- La densificación citoplasmática con filamentos de actina en las adherentes.
- Las placas densas a ambos lados de la membrana con filamentos de citoqueratina divergentes en los desmosomas.
La ultraestructura molecular de cada tipo de unión intercelular, sus proteínas constituyentes y su relevancia en se desarrollan en el Tema 3 · Membrana celular: diferenciaciones y complejos de unión de Biología.
En el pénfigo vulgar, los autoanticuerpos atacan las desmogleínas (proteínas del desmosoma), destruyendo la cohesión entre queratinocitos. El resultado es la acantólisis, es decir, la separación de las células del estrato espinoso que genera ampollas intraepiteliales características. Es un ejemplo directo de la importancia funcional de las máculas adherens.
2.5.3. Diferenciaciones de la superficie basal
La cara basal de las células epiteliales presenta dos tipos de especializaciones.
- Los hemidesmosomas anclan las células a la membrana basal mediante integrinas.
- Los laberintos basales son invaginaciones profundas de la membrana que amplían la superficie de intercambio iónico y se asocian a gran densidad de mitocondrias. Son propios de epitelios con transporte activo intenso, como el túbulo renal o los conductos estriados de glándulas salivales. Al microscopio óptico aparecen como un estriado basal característico.
El detalle ultraestructural de los hemidesmosomas y las integrinas se tratan en Biología de primer curso.
Ver Tema 3 · Membrana celular: diferenciaciones y complejos de unión.
2.5.4. Diferenciaciones de la superficie apical
La cara apical puede desarrollar tres tipos de especializaciones, que se distinguen por tamaño, movilidad y composición del citoesqueleto.
- Las microvellosidades son evaginaciones digitiformes cortas (0,5–1 µm), inmóviles, con citoesqueleto de actina. Su función es aumentar la superficie de absorción. Un enterocito puede llegar a las 3.000 microvellosidades por célula. Al microscopio óptico se reconocen como el borde en cepillo del intestino delgado o del túbulo contorneado proximal.
- Los cilios o cinetocilios son estructuras mucho más largas (7–10 µm) y móviles, con un axonema interno de estructura 9+2 (9 pares de microtúbulos periféricos y 2 centrales) movido por dineínas. Su función es desplazar el contenido luminal (moco en el epitelio respiratorio o el ovocito en la trompa uterina, por ejemplo).
- Los estereocilios son, pese al nombre, microvellosidades largas con citoesqueleto de actina. No tienen axonema y no son móviles. Se encuentran en el epidídimo, donde participan en la absorción del líquido luminal y la maduración espermática, y en las células ciliadas del oído interno como mecanorreceptores.
En el síndrome de Kartagener (discinesia ciliar primaria), las dineínas de los cilios son defectuosas, por lo que los cilios son inmóviles. Esto causa bronquiectasias e infecciones respiratorias recurrentes (aclaramiento mucociliar abolido), infertilidad masculina (espermatozoides inmóviles) y, con frecuencia, situs inversus por alteración del nodo embrionario.
| Diferenciación | Longitud | Citoesqueleto | Movilidad | Función | Ejemplo |
|---|---|---|---|---|---|
| Microvellosidades | 0,5–1 µm | Actina | No | Absorción (↑ superficie) | Enterocito, túbulo proximal |
| Cilios | 7–10 µm | Axonema 9+2 (tubulina) | Sí (batido) | Mover sustancias externas | Tráquea, trompa uterina |
| Estereocilios | Varios µm | Actina | No | Absorción / mecano-recepción | Epidídimo, oído interno |
2.6. Transiciones epiteliales
En la mayoría de las superficies corporales, el cambio de un tipo de epitelio a otro se produce de forma gradual, pasando por formas intermedias. A estas zonas se las llama transiciones epiteliales graduales.
Sin embargo, en determinados puntos anatómicos el cambio de un epitelio a otro es brusco (sin zona de transición). Estas uniones bruscas tienen una gran relevancia clínica porque son zonas de inestabilidad tisular especialmente vulnerables a la metaplasia y la malignización. Los dos ejemplos más relevantes son:
- La unión esofagogástrica: el esófago está revestido por epitelio estratificado plano no queratinizado, el estómago por epitelio simple cilíndrico mucosecretor. La transición es abrupta y visible a simple vista en endoscopia. La exposición crónica del epitelio esofágico al reflujo ácido puede inducir su sustitución por epitelio de tipo columnar intestinal, una metaplasia conocida como esófago de Barrett, con riesgo aumentado de adenocarcinoma.
- La unión escamocolumnar del cérvix uterino: el exocérvix está revestido por epitelio estratificado plano no queratinizado, el endocérvix por epitelio simple cilíndrico mucosecretor. La zona de transición entre ambos es la llamada zona de transformación, localización más frecuente de los cambios displásicos por infección por VPH que preceden al carcinoma de cérvix.
La zona de transformación cervical es la diana principal del cribado citológico (citología de Papanicolaou). La biopsia dirigida por colposcopia de esta zona permite detectar lesiones intraepiteliales de alto grado (CIN 2–3) antes de su progresión a carcinoma invasor.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}