3.1. Diferenciaciones de la membrana plasmática
Las células no tienen membranas uniformes y lisas. En muchos tipos celulares, la membrana plasmática desarrolla especializaciones estructurales adaptadas a funciones concretas: aumentar la superficie de intercambio, reforzar la cohesión con células vecinas o facilitar el transporte de iones. Estas especializaciones son especialmente evidentes en las células epiteliales, que tienen una marcada polaridad: la cara apical, orientada hacia la luz o el exterior, tiene una composición y estructura muy diferente a la cara basolateral.
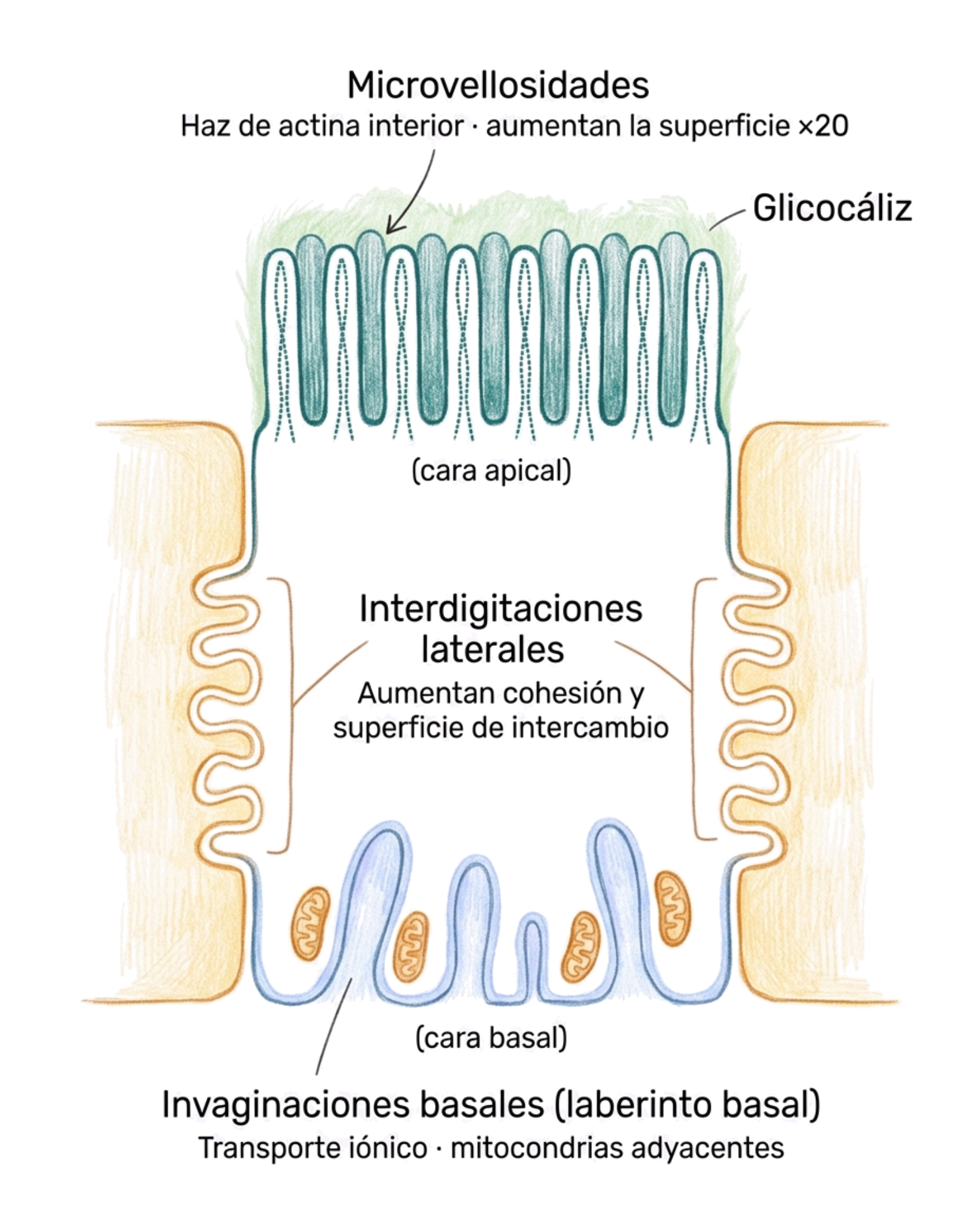
3.1.1. Microvellosidades
Las microvellosidades son extensiones digitiformes de la membrana plasmática apical, presentes principalmente en células dedicadas a la absorción: los enterocitos del intestino delgado y las células del túbulo contorneado proximal del riñón.
Cada microvellosidad mide aproximadamente 1 μm de alto por 0,1 μm de ancho. Son demasiado pequeñas para verse individualmente al microscopio óptico, donde aparecen como una banda densa en la superficie apical denominada ribete en cepillo o borde en chapa. Al microscopio electrónico de transmisión se distinguen perfectamente como estructuras digitiformes individuales.
Una célula intestinal tiene unas 3.000 microvellosidades, lo que aumenta la superficie de absorción aproximadamente 20 veces respecto a una célula sin ellas.
En su interior contienen un haz de 20-30 filamentos de actina paralelos que le dan rigidez y forma. Este haz está estabilizado por proteínas accesorias (villina y fimbrina) y se ancla a la membrana plasmática lateral mediante brazos de miosina I. La membrana que recubre las microvellosidades es especialmente rica en enzimas digestivos y en un glicocáliz muy prominente, lo que facilita la digestión final y la absorción de nutrientes.
La ultraestructura de las microvellosidades y su papel en la absorción intestinal se desarrollan con mayor detalle en Histología, en el contexto del epitelio intestinal.
3.1.2. Interdigitaciones laterales
Las interdigitaciones laterales son entrantes y salientes de la membrana plasmática lateral que encajan con los de la célula vecina, como piezas de un puzle.
Tienen dos funciones principales. Desde el punto de vista mecánico, aumentan la cohesión entre células vecinas al incrementar la superficie de contacto. Desde el punto de vista funcional, aumentan la superficie común entre células adyacentes, facilitando el intercambio de sustancias entre ellas.
Las interdigitaciones no son estructuras pasivas: requieren una concentración adecuada de Ca²⁺ y la participación de los filamentos de actina del citoesqueleto para mantenerse.
3.1.3. Invaginaciones basales
Las invaginaciones basales son pliegues de la membrana plasmática basal que se introducen hacia el interior del citoplasma. Son especialmente prominentes en las células del túbulo contorneado renal, donde forman el denominado laberinto basal.
Su función es aumentar la superficie basolateral disponible para el transporte activo de iones y pequeños solutos. Esto explica la presencia habitual de mitocondrias en posición adyacente a las invaginaciones basales: el transporte activo requiere un aporte continuo de ATP.
3.2. Complejos de unión: visión general
En los tejidos, las células se conectan entre sí mediante estructuras especializadas denominadas complejos de unión. Estas estructuras tienen tres objetivos funcionales distintos:
- Sellar el espacio intercelular para controlar el paso de sustancias entre células (uniones oclusivas).
- Anclar mecánicamente las células entre sí o a la matriz extracelular (uniones de anclaje).
- Comunicar directamente el citoplasma de células adyacentes permitiendo el paso de iones y moléculas pequeñas (uniones comunicantes).
La clasificación clásica distinguía entre zónula (estructura en cinturón que rodea toda la célula) y mácula (estructura puntual, como un botón). La clasificación funcional actual es más útil:
| Función | Tipo | Proteína transmembrana | Citoesqueleto | Ejemplo tisular |
|---|---|---|---|---|
| Oclusiva | Zónula occludens (unión estrecha) | Ocludina · Claudina | Actina (vía ZO-1/2/3) | Epitelio intestinal · epitelio vesical |
| Anclaje célula-célula (actina) | Zónula adherens (unión adherente) | Cadherina E | Actina (vía cateninas) | Epitelios · embrión en desarrollo |
| Anclaje célula-célula (FI) | Desmosoma (mácula adherens) | Desmogleína · Desmocolina | Filamentos intermedios (FI) (vía desmoplaquina) | Piel · miocardio · epitelio vesical |
| Anclaje célula-matriz (FI) | Hemidesmosoma | Integrina α6β4 | Filamentos intermedios (FI) (vía plectina) | Epitelio basal · piel |
| Comunicante | Unión GAP | Conexina (×6 = conexón) | No aplica | Miocardio · músculo liso · hepatocitos · SNC |
Especializaciones de la membrana plasmática que conectan células adyacentes con tres funciones: sellado del espacio intercelular (uniones oclusivas), anclaje mecánico (uniones de anclaje) y comunicación directa entre citoplasmas (uniones comunicantes).
3.3. Uniones oclusivas: la zónula occludens
La zónula occludens o unión estrecha es la primera unión que encontramos al descender desde la superficie apical de las células epiteliales. Se extiende en forma de cinturón continuo alrededor de toda la célula, sellando completamente el espacio intercelular en esa zona.
Estructuralmente, las proteínas transmembrana de células vecinas contactan directamente entre sí, eliminando el espacio intercelular en ese punto. Al microscopio electrónico, la zona de contacto aparece como una fusión de las membranas. Mediante criofractura se observan hileras paralelas de proteínas transmembrana formando un patrón de «pespunte».
Las proteínas transmembrana principales son ocludina y claudina. Estas proteínas se anclan en el citoplasma a proteínas periféricas internas denominadas proteínas ZO (Zónula Occludens 1, 2 y 3), que a su vez se unen al citoesqueleto de actina.
La función de la zónula occludens es doble:
- Barrera paracelular: limita el paso de sustancias a través del espacio entre células (vía paracelular). El sello no es absoluto: permite el paso selectivo de iones, monosacáridos y aminoácidos en función del número y tipo de claudinas presentes. Las células intestinales son más permeables al Na⁺ que las células de la vejiga.
- Barrera de difusión lateral: impide que los lípidos y proteínas de la superficie apical difundan hacia la superficie basolateral y viceversa, manteniendo la polaridad de la célula epitelial.
Las claudinas son esenciales para la reabsorción de iones específicos. Las mutaciones en la claudina-16 producen hipomagnesemia familiar con hipercalciuria y nefrocalcinosis: sin claudina-16 funcional, el Mg²⁺ no puede reabsorberse en el asa de Henle y se pierde por la orina.
3.4. Uniones de anclaje
Las uniones de anclaje conectan el citoesqueleto de células adyacentes o de la célula con la matriz extracelular, transmitiendo fuerzas mecánicas y confiriendo resistencia a los tejidos. Tienen siempre dos componentes: una proteína transmembrana de anclaje (cadherina o integrina) y una placa interna de anclaje en el citoplasma que conecta con el citoesqueleto.
3.4.1. Uniones adherentes
La unión adherente o zónula adherens es una unión en cinturón que rodea la célula por debajo de la zónula occludens. Une células adyacentes mediante cadherinas (proteínas transmembrana calcio-dependientes) que contactan con las cadherinas de la célula vecina.
En el citoplasma, las cadherinas se unen a cateninas, que a su vez se anclan a los filamentos de actina del citoesqueleto. Esta conexión actina-catenina-cadherina forma un cinturón contráctil que tiene una función especialmente relevante durante el desarrollo embrionario, donde los cambios en la tensión de este cinturón permiten el plegamiento de capas epiteliales.
3.4.2. Desmosomas
El desmosoma o mácula adherens es una unión puntual (en botón) que ancla células adyacentes mediante una placa densa en el citoplasma. A diferencia de las uniones adherentes, los desmosomas utilizan filamentos intermedios en lugar de actina.
Su estructura incluye proteínas transmembrana de la familia de las cadherinas: desmogleína y desmocolina. Estas se unen en el citoplasma a proteínas de la placa interna: desmoplaquina y placoglobina, que anclan los filamentos intermedios (queratina en células epiteliales, desmina en células musculares).
Los desmosomas son especialmente abundantes en tejidos sometidos a estrés mecánico intenso: epitelio de la piel, miocardio y epitelio vesical.
El pénfigo vulgar es una enfermedad autoinmune en la que el paciente produce anticuerpos contra la desmogleína 3, una de las cadherinas del desmosoma. La pérdida de función de los desmosomas provoca la separación de las células epidérmicas (acantólisis), formación de ampollas intraepidérmicas con pérdida masiva de líquido. Es una enfermedad grave que puede ser mortal sin tratamiento.
3.4.3. Hemidesmosomas
Los hemidesmosomas son estructuras similares a medio desmosoma que anclan la célula a la membrana basal y a la matriz extracelular subyacente. A diferencia de los desmosomas, utilizan integrinas como proteínas transmembrana (no cadherinas) y se unen a la laminina de la membrana basal en el exterior.
En el citoplasma, las integrinas se conectan a la plectina y a la BP230, que anclan los filamentos intermedios de queratina. Los hemidesmosomas son esenciales para fijar el epitelio a su base.
| Característica | Desmosoma | Hemidesmosoma |
|---|---|---|
| Tipo de unión | Célula a célula | Célula a matriz extracelular |
| Proteína transmembrana | Desmogleína y desmocolina (cadherinas) | Integrina α6β4 |
| Proteínas de placa interna | Desmoplaquina · placoglobina | Plectina · BP230 |
| Citoesqueleto | Filamentos intermedios (queratina en epitelio, desmina en músculo) | Filamentos intermedios de queratina |
| Contraparte exterior | Cadherinas de la célula vecina | Laminina de la membrana basal |
| Morfología | Mácula (botón puntual) — espacio intercelular aumentado | Semibotón en la cara basal |
| Patología asociada | Pénfigo vulgar (anti-desmogleína 3) | Penfigoide ampolloso (anti-BP180/BP230) |
3.5. Uniones comunicantes: las uniones GAP
Las uniones GAP o uniones comunicantes son canales intercelulares que conectan directamente el citoplasma de dos células adyacentes, permitiendo el paso de iones y moléculas pequeñas sin necesidad de cruzar ninguna membrana.
Como se describió en el contexto de las sinapsis eléctricas, cada canal está formado por dos conexones enfrentados, uno aportado por cada célula. Cada conexón está compuesto por seis conexinas, proteínas transmembrana de ~25 kDa. El canal resultante permite el paso de moléculas de hasta ~1.000 Da: iones, AMPc, IP₃, glucosa y otros metabolitos pequeños.
Las uniones GAP son especialmente abundantes en:
- Miocardio: sincronizan la contracción de los cardiomiocitos, permitiendo que el corazón funcione como un sincitio funcional.
- Músculo liso visceral: coordinan las ondas de contracción peristáltica del intestino y del útero.
- Hepatocitos: permiten la coordinación metabólica entre células hepáticas adyacentes.
- Sistema nervioso: en algunos circuitos del SNC y SNP, donde actúan como sinapsis eléctricas.
Las uniones GAP como base de las sinapsis eléctricas, la regulación de los conexones por Ca²⁺ y H⁺, y su papel en la sincronización del miocardio se desarrollan en T15 de Fisiología — Introducción al estudio de las sinapsis.
