16.1. El cromosoma interfásico
El cromosoma metafásico, compacto y visible al microscopio óptico, es la forma minoritaria: la célula solo lo adopta durante unas pocas horas en cada ciclo. La mayor parte del tiempo, el cromosoma existe en forma de cromatina interfásica, descondensada, transcripcionalmente activa e invisible como estructura individual al microscopio óptico convencional.
La transición entre los dos estados es reversible y cíclica. Al entrar en mitosis, la cromatina se condensa progresivamente hasta formar el cromosoma metafásico. Al salir de la mitosis, en telofase, los cromosomas se descondensan y recuperan la organización interfásica. Este proceso se repite en cada división celular.
Descondensado no significa desorganizado. Dentro del núcleo interfásico, cada cromosoma ocupa una región espacial delimitada, no mezcla su cromatina con la de los cromosomas vecinos y mantiene una posición preferente dentro del núcleo. Esta organización tiene consecuencias funcionales directas.
16.2. Eucromatina y heterocromatina
16.2.1. Definición y propiedades
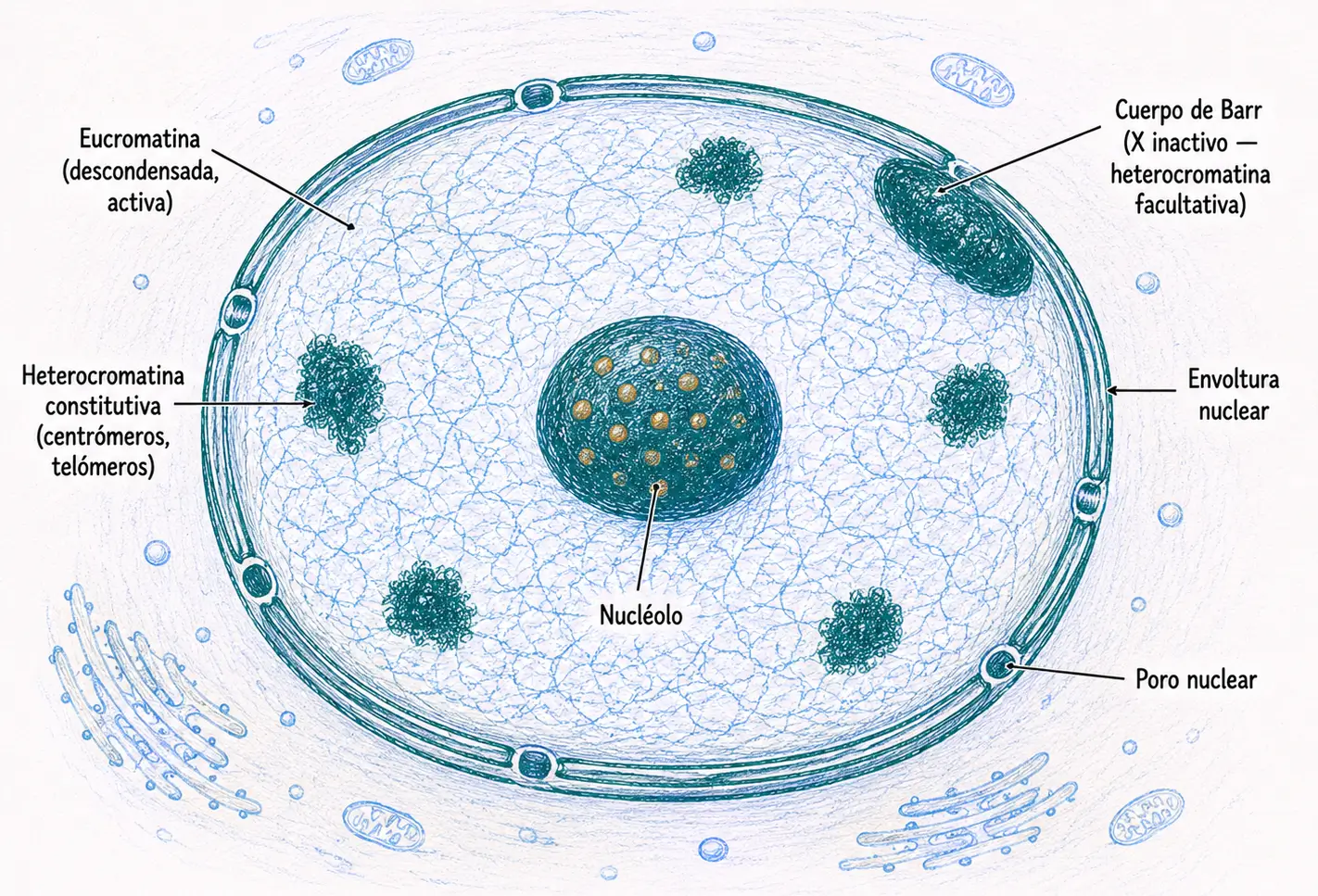
El núcleo interfásico no es homogéneo. Cuando se tiñe con colorantes básicos como la hematoxilina, aparecen dos tipos de regiones con intensidad diferente: zonas densamente teñidas, donde la cromatina está más compactada, y zonas claras, donde está más laxa. Esta diferencia morfológica refleja diferencias funcionales profundas y da lugar a la distinción entre eucromatina y heterocromatina.
Eucromatina: regiones de la cromatina que permanecen descondensadas en interfase, se tiñen débilmente con colorantes básicos y contienen genes transcritos activamente.
Heterocromatina: regiones que permanecen condensadas en interfase, se tiñen intensamente y corresponden a zonas transcripcionalmente inactivas o con baja actividad génica.
La diferencia entre ambas no es estática ni absoluta. Una región puede pasar de eucromatina a heterocromatina y viceversa en función del tipo celular, del momento del desarrollo o de señales reguladoras. Los mecanismos moleculares que gobiernan esta transición, basados principalmente en modificaciones de las colas de histonas y en la metilación del ADN, pertenecen al campo de la epigenética.
Los mecanismos moleculares que convierten eucromatina en heterocromatina y viceversa (acetilación/desacetilación de histonas, metilación del ADN, proteínas del grupo Polycomb) se estudian en Bioquímica y Biología Molecular.
16.2.2. Heterocromatina constitutiva y facultativa
Dentro de la heterocromatina se distinguen dos categorías según su comportamiento en distintos tipos celulares.
- La heterocromatina constitutiva está condensada e inactiva en todas las células del organismo, independientemente del tipo celular o del estadio del desarrollo. No contiene genes funcionalmente relevantes, sino secuencias repetitivas altamente organizadas. Se localiza principalmente en los centrómeros, los telómeros, los satélites de los cromosomas acrocéntricos y algunas regiones pericentroméricas. Su función es estructural: mantiene la integridad de las regiones críticas del cromosoma.
- La heterocromatina facultativa, en cambio, contiene genes que son activos en algunos tipos celulares pero inactivos en otros. En los tipos celulares donde esos genes deben silenciarse, la cromatina correspondiente se condensa y adopta el aspecto de heterocromatina. El ejemplo más claro y clínicamente relevante en biología humana es la inactivación del cromosoma X.
La heterocromatina constitutiva es siempre heterocromatina, en cualquier célula. La heterocromatina facultativa puede ser eucromatina en una célula y heterocromatina en otra, Es la misma secuencia de ADN, con actividad diferente según el contexto celular.
16.2.3. El cuerpo de Barr
Las mujeres tienen dos cromosomas X en cada célula somática, pero solo uno está activo en cada célula concreta. El otro queda completamente silenciado. Se condensa como heterocromatina facultativa y forma el cuerpo de Barr (también llamado corpúsculo sexual de Barr o cromatina sexual). Este proceso se denomina inactivación del X o lionización, en honor a Mary Lyon, quien lo describió en 1961.
El cuerpo de Barr es visible al microscopio óptico como un corpúsculo denso adherido a la cara interna de la envoltura nuclear, de aproximadamente 1 µm de diámetro. En las células de la mucosa oral es especialmente fácil de identificar con tinción de hematoxilina-eosina o con orceína.
El número de cuerpos de Barr en una célula es igual al número de cromosomas X menos uno: n(cuerpos de Barr) = n(X) − 1. Una mujer 46,XX tiene 1 cuerpo de Barr. Un hombre 46,XY tiene 0. Una persona con síndrome de Klinefelter (47,XXY) tiene 1. Una mujer 47,XXX tiene 2.
La inactivación del cromosoma X es aleatoria. En cada célula, el X inactivado puede ser el de origen materno o el de origen paterno, y esa elección se mantiene de forma estable en todas las células hijas. Esto explica el mosaicismo de expresión característico de las mujeres portadoras de enfermedades ligadas al X: dependiendo de qué proporción de células inactivan el X patológico o el normal, la expresión clínica varía.
El cuerpo de Barr tuvo aplicación diagnóstica histórica en citogenética clínica: contar los corpúsculos de Barr en células de mucosa oral permitía estimar el número de cromosomas X antes de la disponibilidad generalizada del cariotipo. Hoy se usa el cariotipo y la FISH, pero el concepto sigue siendo relevante para entender enfermedades como el síndrome de Turner (45,X: 0 cuerpos de Barr) o el síndrome de Klinefelter (47,XXY: 1 cuerpo de Barr).
16.3. Organización espacial: territorios cromosómicos
Durante décadas se asumió que los cromosomas interfásicos se distribuían al azar dentro del núcleo, mezclando su cromatina en un espacio compartido. Las técnicas de hibridación in situ fluorescente (FISH) con sondas cromosoma-específicas demostraron que esto no es así.
Cada cromosoma ocupa en interfase una región nuclear discreta, compacta y no entremezclada con la cromatina de los cromosomas vecinos. Estas regiones se denominan territorios cromosómicos. Los experimentos de FISH muestran que los territorios son posiciones preferentes y reproducibles entre células del mismo tipo, no posiciones aleatorias.
La localización dentro del núcleo no es trivial. Los cromosomas con mayor densidad génica y mayor actividad transcripcional tienden a ocupar el interior del núcleo, donde el acceso a la maquinaria de transcripción es mayor. Los cromosomas más silenciados, incluido el X inactivo, tienden a localizarse en la periferia nuclear, próximos a la lámina nuclear que, como se vio en el tema 13, actúa como plataforma de anclaje para regiones de cromatina inactiva.
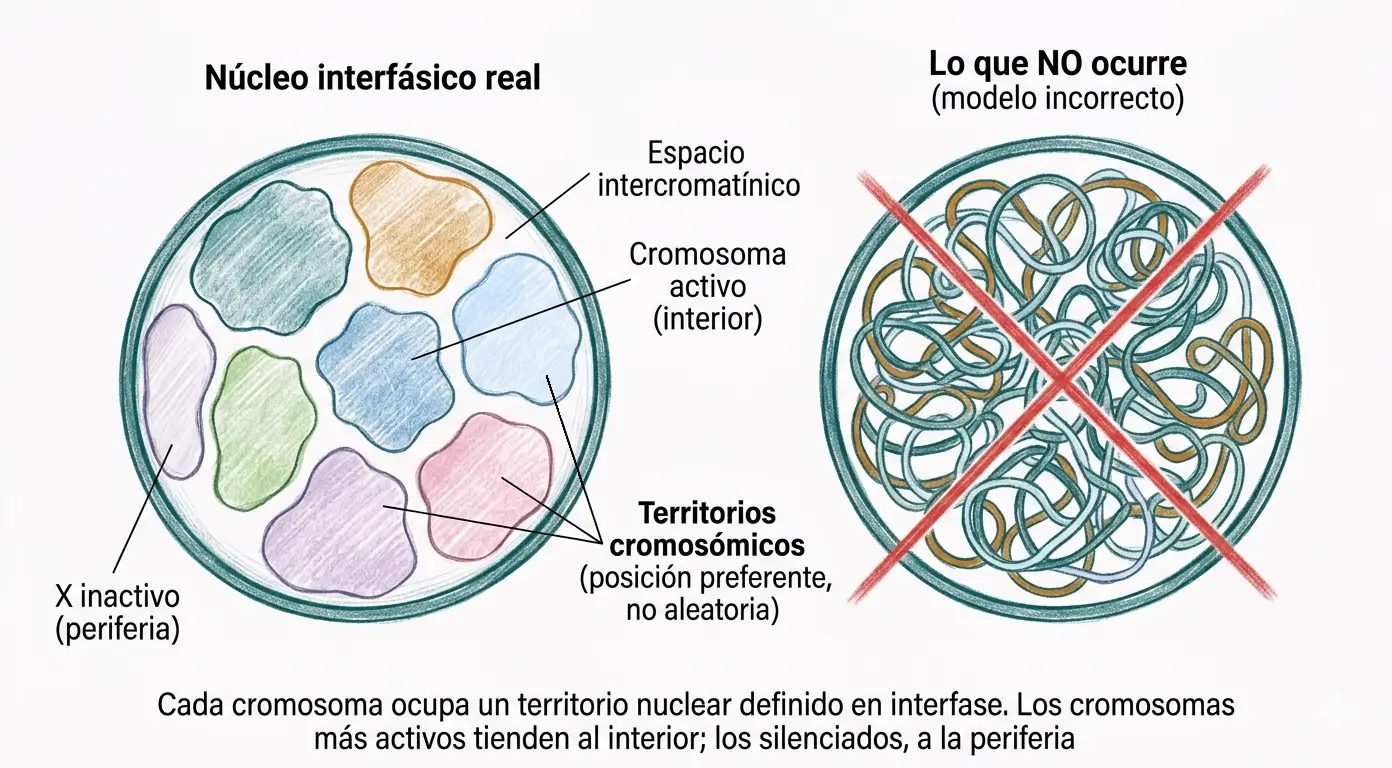
Los territorios cromosómicos no son compartimentos completamente estancos. Existe un espacio intercromatínico entre territorios por donde circulan factores de transcripción, ARN mensajero en procesamiento y complejos de reparación del ADN. Los genes situados en los límites de un territorio pueden proyectarse hacia el espacio intercromatínico para facilitar su transcripción.
16.4. El ciclo del cromosoma
16.4.1. Estado del cromosoma según la fase del ciclo
El estado del cromosoma cambia de forma coordinada con las fases del ciclo celular. Esos cambios afectan al grado de condensación, al número de cromátidas y a la actividad transcripcional.
| Fase del ciclo | Estado de condensación | Cromátidas/cromosoma | Actividad transcripcional |
|---|---|---|---|
| G1 | Descondensado (cromatina interfásica) | 1 | Alta |
| S | Descondensado (replicación en curso) | 1 → 2 | Alta (salvo regiones en replicación activa) |
| G2 | Descondensado / inicio de condensación | 2 | Alta |
| Profase | Condensación progresiva | 2 | Activa (decrece al final) |
| Metafase / Anafase | Máxima condensación | 2 / 1 | Nula |
| Telofase | Descondensación progresiva | 1 | Se reanuda |
16.4.2. Actividad transcripcional a lo largo del ciclo
La transcripción requiere acceso de la ARN polimerasa y de los factores de transcripción al ADN. Ese acceso depende directamente del grado de compactación de la cromatina. Cuando el cromosoma está en su forma máximamente condensada, el ADN es prácticamente inaccesible y la transcripción se detiene.
En interfase, la cromatina está suficientemente descondensada como para que los factores de transcripción accedan a los promotores de los genes activos. No toda la cromatina interfásica se transcribe por igual. Las regiones de eucromatina son las más accesibles, mientras que las regiones de heterocromatina permanecen silenciosas incluso en interfase.
La condensación máxima del cromosoma en metafase tiene un coste funcional. La transcripción se detiene completamente, por eso la profase tardía ya muestra una caída de la actividad transcripcional y la reanudación en telofase temprana es uno de los primeros eventos de la reconstrucción del núcleo hijo.
Un indicador sensible de esta reanudación es la reaparición del nucléolo. El nucléolo desaparece en profase porque el ADN ribosómico queda condensado en los cromosomas acrocéntricos. En telofase, cuando esos cromosomas se descondensan y se reconstituye la envoltura nuclear, la transcripción del ADNr se reanuda y los nucléolos vuelven a aparecer: primero múltiples y pequeños, uno por cada organizador nucleolar activo, y luego se fusionan hasta formar 1-2 nucléolos de mayor tamaño.
