19.1. La división celular: necesidad y regulación
Todas las células de un organismo están destinadas a morir. Para mantener la homeostasis tisular, cada célula que muere debe ser reemplazada por un número equivalente de células nuevas. Este equilibrio no es trivial: si la tasa de división es insuficiente, el tejido pierde células progresivamente y puede desarrollar aplasia. Si la tasa es excesiva, el tejido acumula células de forma descontrolada, lo que puede conducir a procesos neoplásicos.
La división celular debe estar regulada con precisión. La desregulación en uno u otro sentido (escasa o aumentada) tiene consecuencias patológicas directas.
Antes de dividirse, la célula debe prepararse duplicando su masa, sus orgánulos y, sobre todo, su material genético. La duplicación del ADN debe ser exacta. El error en la segregación cromosómica es de aproximadamente 1 por cada 100.000 divisiones. Cualquier error en el reparto cromosómico puede generar células con un número anómalo de cromosomas, lo que se denomina aneuploidía.
La división celular comprende dos procesos distintos que ocurren de forma coordinada:
- La mitosis, que es el reparto del material cromosómico entre dos núcleos hijos.
- La citocinesis, que es la división del citoplasma en dos células hijas independientes.
19.1.1. Tipos de mitosis
En eucariotas existen dos modalidades de mitosis según el comportamiento de la envoltura nuclear.
- Mitosis abierta: propia de animales superiores, la envoltura nuclear se desensambla al inicio de la división y se reconstituye al final.
- Mitosis cerrada: la realizan algunos protozoos y hongos, los cromosomas se segregan sin que la membrana nuclear llegue a romperse.
19.1.2. Inicio de la fase M: papel de las CDK
La entrada en mitosis está desencadenada por las proteínas quinasas dependientes de ciclina (CDK), que fosforilan un conjunto de sustratos proteicos cuyos cambios son los eventos más visibles del inicio de la fase M:
- La fosforilación de las laminas nucleares provoca la desestructuración de la lámina nuclear y, con ella, la ruptura de la envoltura nuclear.
- La fosforilación de las condensinas desencadena la condensación progresiva de los cromosomas.
- El retículo endoplásmico y el aparato de Golgi se fragmentan y redistribuyen.
- La célula pierde sus conexiones con la matriz extracelular y con las células vecinas.
- El citoesqueleto interfásico se reorganiza para construir la maquinaria mitótica.
19.2. Preparación del cromosoma para la división: condensinas y cohesinas
Antes de que comience la mitosis propiamente dicha, los cromosomas deben estar correctamente preparados. Dos familias de proteínas con estructura similar son responsables de este proceso: las condensinas y las cohesinas.
- Las condensinas son complejos proteicos que utilizan la energía de la hidrólisis de ATP para enrollar el ADN e inducir la compactación progresiva de cada cromátida alrededor de un eje central lineal. Este proceso genera bucles de cromatina que compactan la cromátida de forma ordenada a lo largo de toda su longitud.
- Las cohesinas se depositan entre las dos cromátidas hermanas durante la replicación del ADN en fase S y las mantienen estrechamente unidas hasta la anafase. Su estructura, similar a la de las condensinas, forma un anillo que abraza a las dos cromátidas.
Condensinas y cohesinas no actúan de forma independiente: para que las condensinas puedan compactar correctamente una cromátida, las cohesinas deben mantener las dos cromátidas hermanas unidas y alineadas. Solo así se evitan errores de segregación.
A medida que avanza la condensación en profase, las cohesinas de los brazos cromosómicos se van liberando progresivamente, lo que permite visualizar las dos cromátidas de cada cromosoma. Sin embargo, las cohesinas de la región centromérica persisten hasta la anafase, donde serán escindidas por la separasa para permitir la segregación.
19.3. Duplicación del centrosoma
El centrosoma, formado por un par de centríolos rodeados de matriz pericentriolar, es el principal centro organizador de microtúbulos (MTOC) de la célula animal. Debe duplicarse antes de la mitosis para que se puedan formar los dos polos del huso.
- La duplicación del centrosoma comienza en G1, cuando los dos centríolos del par se separan ligeramente.
- Durante la fase S, perpendicular a la base de cada centríolo parental comienza a formarse un centríolo hijo, de forma simultánea a la replicación del ADN.
- El centríolo hijo completa su crecimiento en G2. Los dos pares de centríolos permanecen juntos formando un único complejo centrosómico hasta el inicio de la fase M.
- En la fase M, los centríolos se separan y migran hacia polos opuestos de la célula, donde nuclearán los dos ásteres del huso mitótico.
Al finalizar la división, cada célula hija recibe un centrosoma con un par de centríolos: un centríolo parental y uno hijo. El centríolo hijo madurará durante el siguiente ciclo y se replicará a su vez, restaurando la situación inicial.
19.4. Fases de la mitosis
La mitosis se divide en cinco fases: profase, prometafase, metafase, anafase y telofase. La más larga, paradójicamente, es la metafase.
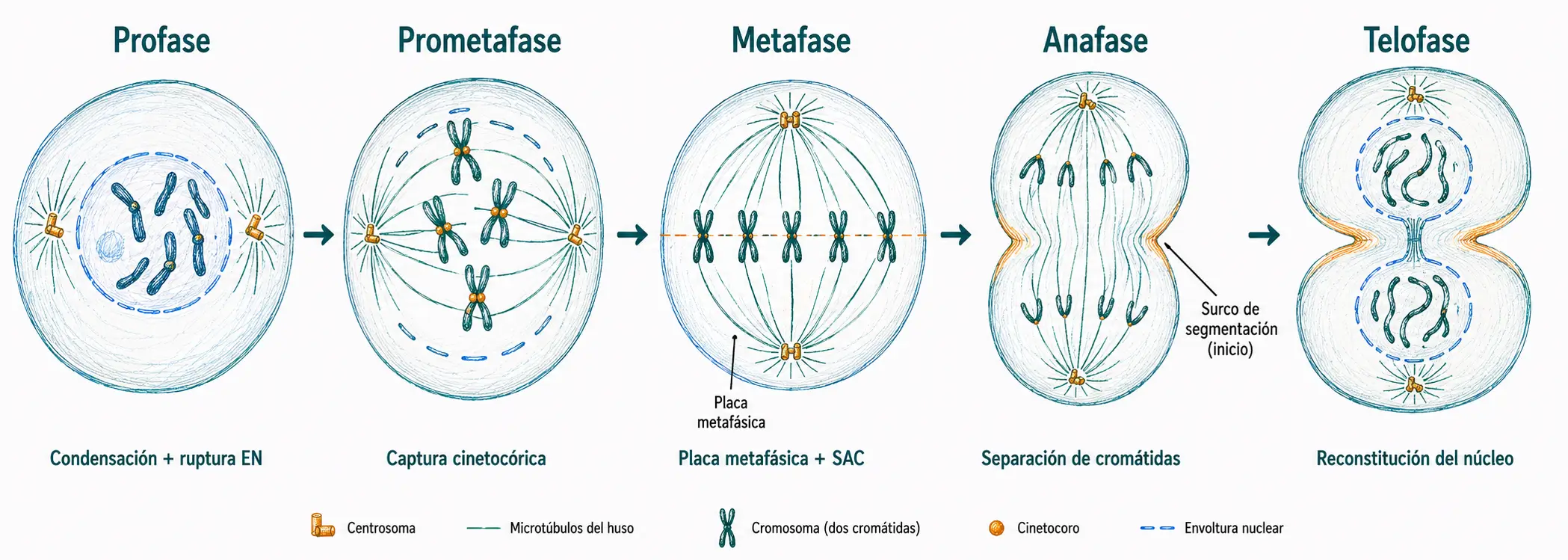
19.4.1. Profase
La profase se define desde el momento en que comienza la condensación cromosómica hasta el instante anterior a la ruptura de la envoltura nuclear, que permanece intacta durante toda esta fase.
Los cromosomas se condensan progresivamente. Hacia la mitad de la profase ya son visibles las dos cromátidas de cada cromosoma, separadas excepto en la región del centrómero, donde las cohesinas centroméricas las mantienen unidas. En esa región centromérica comienzan a esbozarse los cinetocoros, estructuras proteicas que servirán de punto de anclaje para los microtúbulos del huso. El nucléolo desaparece porque el ADN organizador nucleolar queda condensado en el cromosoma, impidiendo la transcripción del ARNr.
En el citoplasma, los microtúbulos interfásicos se despolimerizan: las proteínas catastrofinas son fosforiladas por las CDK, lo que aumenta la frecuencia de catástrofes microtubulares. Simultáneamente, cada centrosoma nuclea un nuevo conjunto de microtúbulos radiales muy dinámicos (vida media de segundos) que forman el áster. Los pares de centríolos comienzan a migrar hacia polos opuestos de la célula.
19.4.2. Prometafase
La prometafase comienza con la ruptura de la envoltura nuclear, cuyos fragmentos se dispersan como vesículas del retículo endoplásmico. Este evento da acceso a los microtúbulos del huso a los cromosomas condensados.
En esta fase se completa la diferenciación de los cinetocoros. Los cinetocoros son dos placas proteicas trilaminares (en mamíferos) situadas una a cada lado del centrómero, enfrentadas entre sí. En la especie humana, cada cinetocoro se une a unos 40 microtúbulos cinetocóricos.
La captura de los cromosomas por el huso sigue un mecanismo de «busca y captura». Los microtúbulos del áster crecen dinámicamente hacia el interior de la célula hasta contactar lateralmente con un cinetocoro. El cinetocoro se desliza entonces a lo largo del microtúbulo hacia el polo, y la adhesión lateral se convierte en adhesión terminal. Al mismo tiempo, microtúbulos del polo opuesto alcanzan el segundo cinetocoro del mismo cromosoma, estableciendo la adhesión bipolar: cada cromosoma queda unido a ambos polos del huso, con un cinetocoro orientado hacia cada polo.
Una vez establecida la adhesión bipolar, el cromosoma experimenta fuerzas opuestas que lo desplazan hacia el centro de la célula. La prometafase termina cuando todos los cromosomas han alcanzado el plano ecuatorial con adhesión bipolar completa.
El huso mitótico completo contiene tres tipos de microtúbulos con funciones distintas:
- Los microtúbulos astrales irradian desde cada centrosoma hacia la corteza celular y posicionan el huso en el centro de la célula.
- Los microtúbulos cinetocóricos se extienden desde cada polo hasta los cinetocoros de los cromosomas y son los responsables directos de la segregación.
- Los microtúbulos polares (o solapados) nacen en los polos y se solapan en la zona ecuatorial con los del polo opuesto; proteínas motoras los empujan entre sí, contribuyendo a separar los polos y alargar el huso.
19.4.3. Metafase
En metafase, todos los cromosomas están alineados en el plano ecuatorial formando la placa metafásica. Es el momento de máxima condensación cromosómica y de mínima movilidad aparente. Sin embargo, los cromosomas no están completamente inmóviles, realizan pequeñas oscilaciones generadas por la tensión entre las fuerzas que tiran de ellos hacia cada polo.
La metafase es, sorprendentemente, la fase de mayor duración de la mitosis. Esta duración no es casual: responde a un mecanismo de seguridad denominado punto de control del ensamblaje del huso (SAC, del inglés spindle assembly checkpoint).
El SAC detecta cualquier cinetocoro que no haya logrado adhesión bipolar y mantiene inhibida la transición metafase-anafase hasta que todos los cromosomas estén correctamente unidos al huso. Basta con que un solo cinetocoro esté libre para detener la mitosis. Este mecanismo evita la segregación asimétrica y la aneuploidía.
Las proteínas clave del SAC, como Mad2 y BubR1, se acumulan en los cinetocoros libres y generan una señal inhibitoria que bloquea el Complejo Promotor de la Anafase (APC). Solo cuando todos los cinetocoros están bajo tensión bipolar desaparece esta señal y el APC puede activarse, desencadenando la anafase.
19.4.4. Anafase
La anafase comienza cuando la célula ha verificado que todos los cromosomas tienen adhesión bipolar completa. Es la fase más breve de la mitosis, y en ella ocurren dos cosas simultáneas: la separación de las cromátidas hermanas y el alargamiento general del huso.
Mecanismo de separación de cromátidas:
La señal de entrada en anafase es la activación del Complejo Promotor de la Anafase (APC), una ubiquitín-ligasa que ubiquitina la proteína segurina. La segurina, mientras está intacta, actúa como inhibidor de la separasa. Al ser degradada por el APC, la separasa queda libre y escinde la subunidad Scc1 del complejo de cohesinas centroméricas. Sin cohesinas, las dos cromátidas hermanas se separan bruscamente y son arrastradas hacia polos opuestos.
Cascada de separación en anafase:
APC activado → ubiquitinación de segurina → degradación de segurina → liberación de separasa → escisión de cohesinas centroméricas → separación de cromátidas hermanas.
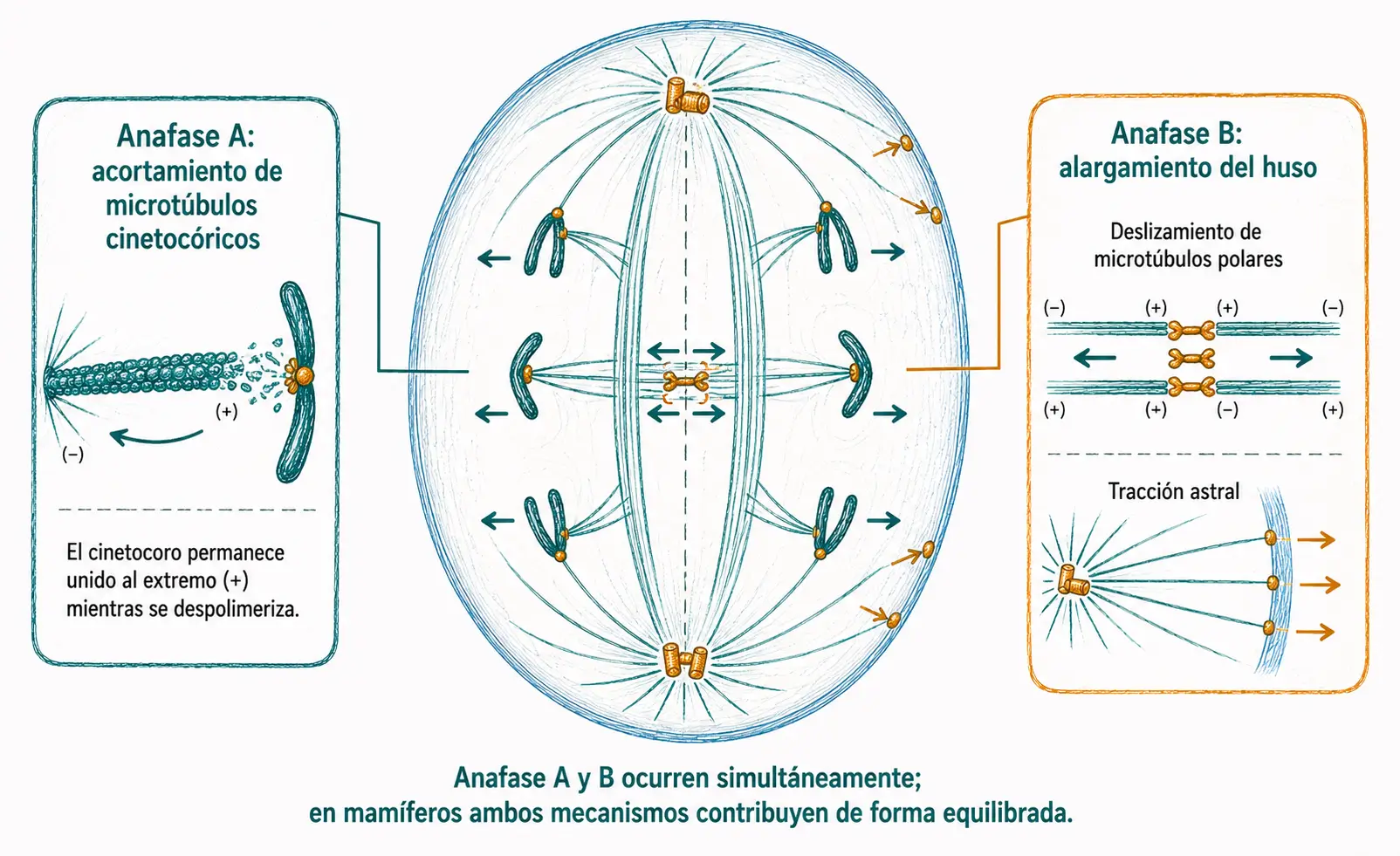
Anafase A y anafase B:
El movimiento cromosómico hacia los polos está generado por dos mecanismos complementarios que ocurren simultáneamente.
- En la anafase A, los microtúbulos cinetocóricos se acortan por despolimerización del extremo (+) en el punto de unión con el cinetocoro. El cinetocoro permanece unido al microtúbulo mientras este se desensambla, siendo arrastrado hacia el polo. Las proteínas motoras del cinetocoro contribuyen a mantener la unión durante el proceso.
- En la anafase B, el huso se alarga por dos fuerzas independientes. Las proteínas motoras dirigidas hacia el extremo (+) entrecruzan los microtúbulos polares antiparalelos de ambos semihijos y los deslizan entre sí, separando los polos. Simultáneamente, proteínas motoras dirigidas hacia el extremo (-) se anclan al córtex celular y tiran de los microtúbulos astrales hacia fuera, alejando los polos de la célula.
En mamíferos ambos mecanismos contribuyen de forma equilibrada; en otros organismos predomina uno de ellos.
Simultáneamente al movimiento cromosómico, en el plano ecuatorial comienzan a ensamblarse los filamentos de actina y miosina II que formarán el anillo contráctil de la citocinesis. En células animales ya es visible una ligera invaginación de la membrana plasmática en la zona ecuatorial.
19.4.5. Telofase
La telofase comienza cuando los dos lotes de cromosomas hijos han llegado a los polos celulares. Los microtúbulos cinetocóricos desaparecen; solo persisten los microtúbulos polares, que forman un haz paralelo entre los dos núcleos en formación y marcan el eje de la futura citocinesis.
La envoltura nuclear se reconstituye:
- Pequeñas vesículas del retículo endoplásmico comienzan a fusionarse alrededor de los cromosomas, formando primero grupos de 2-3 cromosomas envueltos, que se fusionan progresivamente hasta formar un núcleo único.
- Las laminas nucleares, que estaban fosforiladas durante la mitosis, se defosforilan y reconstituyen la lámina densa.
- Los complejos del poro nuclear aparecen precozmente para controlar el tráfico hacia el nuevo núcleo, que aumenta de tamaño por entrada de agua y proteínas nucleares.
En el nuevo núcleo se reanuda la actividad metabólica:
- Se inicia la transcripción y reaparecen los nucléolos, primero pequeños y numerosos, que se fusionan hasta formar 1-2 nucléolos grandes.
- Los cromosomas se descondensan progresivamente hasta adoptar el aspecto de cromatina interfásica.
19.5. Citocinesis
La citocinesis es la división del citoplasma que completa la separación en dos células hijas. Comienza durante la anafase y termina en la telofase tardía.
Al entrar en mitosis, la red cortical de actina y miosina II interfásica se desensambla. Cuando las cromátidas se separan en anafase, la miosina II comienza a acumularse en el plano ecuatorial, ensamblando el anillo contráctil: una banda circunferencial de filamentos de actina y miosina II dispuesta justo por debajo de la membrana plasmática.
La posición del anillo está determinada por los microtúbulos polares: el haz de microtúbulos solapados antiparalelos que persiste en el plano ecuatorial durante la telofase actúa como señal espacial que marca dónde debe contraerse la célula.
La miosina II contrae el anillo progresivamente, estrangulando la célula y formando el surco de segmentación. La contracción continúa hasta que la membrana plasmática de los dos lados contacta, dejando las dos células unidas únicamente por un delgado puente citoplasmático denominado cuerpo medio, que contiene los restos de los microtúbulos polares fuertemente empaquetados. Finalmente el cuerpo medio se rompe, completando la separación.
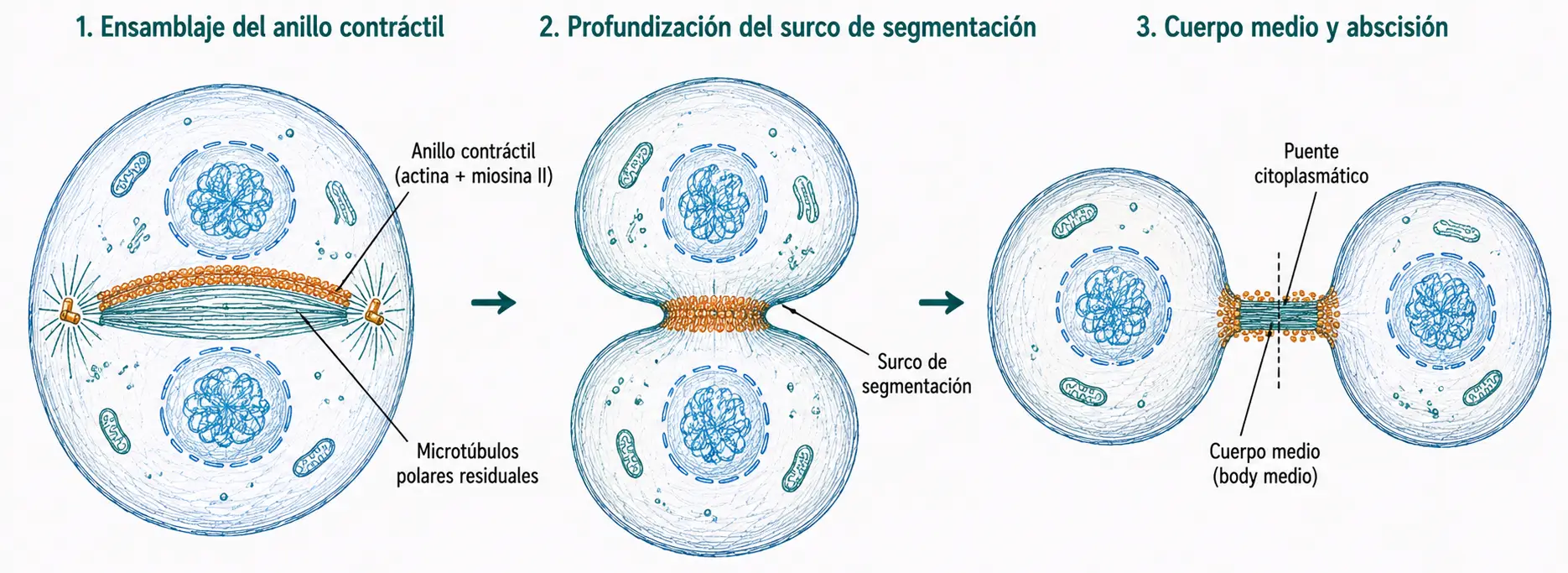
En células animales la citocinesis es simétrica y se produce por el plano ecuatorial. En células vegetales no existe anillo contráctil: la división se produce por síntesis de una nueva pared celular (fragmoplasto) en el plano ecuatorial, un mecanismo completamente distinto.
19.6. Evolución del material genético durante la mitosis
A lo largo del ciclo celular, la cantidad de ADN y el número de cromátidas por cromosoma varían de forma predecible. La ploidía (número de juegos cromosómicos) se mantiene constante en 2n durante toda la mitosis.
| Fase | Ploidía | Cromátidas/cromosoma | Cantidad de ADN |
|---|---|---|---|
| G1 | 2n | 1 | 2C |
| S / G2 | 2n | 2 | 4C |
| Profase – Metafase | 2n | 2 | 4C |
| Anafase | 4n* | 1 | 4C |
| Telofase / células hijas | 2n | 1 | 2C |
Durante la anafase, mientras las cromátidas se están desplazando hacia los polos pero antes de completarse la citocinesis, la célula contiene transitoriamente 4n cromosomas individuales. Al terminar la citocinesis, cada célula hija recupera la dotación 2n con 2C de ADN.
La ploidía no cambia durante la mitosis: las células hijas tienen el mismo número de cromosomas que la célula madre (2n). Lo que cambia es la cantidad de ADN por cromatidio y el número de cromátidas por cromosoma, no el número de juegos cromosómicos.
19.7. Relevancia clínica: antimitóticos y aneuploidía
Fármacos antimitóticos en oncología
Varios fármacos de uso clínico actúan sobre el huso mitótico. Los alcaloides de la vinca (vincristina, vinblastina) inhiben la polimerización de la tubulina, impidiendo la formación del huso y bloqueando la mitosis en prometafase. Los taxanos (paclitaxel, docetaxel) estabilizan los microtúbulos impidiendo su despolimerización, lo que bloquea igualmente la dinámica del huso y detiene la mitosis. En ambos casos, la activación prolongada del SAC lleva a la célula a la apoptosis. Estos fármacos son especialmente eficaces en células con alta tasa de división, como las tumorales, aunque también afectan a tejidos de renovación rápida (médula ósea, epitelio intestinal), lo que explica sus efectos secundarios.
Aneuploidía: errores de segregación
Los fallos en el SAC o en la adhesión bipolar pueden producir aneuploidías, es decir, células hijas con un número anómalo de cromosomas. La trisomía 21 (síndrome de Down) es el ejemplo más conocido: resulta de la no disyunción del cromosoma 21 durante la meiosis, pero los mecanismos de error son similares en mitosis. En las células tumorales, la inestabilidad cromosómica, frecuentemente asociada a defectos del SAC, genera aneuploidías complejas que contribuyen a la progresión del cáncer.
