5.1. Funciones generales de la membrana plasmática
La membrana plasmática no es una barrera pasiva. Además de delimitar la célula y mantener la diferencia entre el medio intracelular y el extracelular, realiza funciones activas esenciales para la supervivencia y coordinación celular.
Sus funciones principales son:
- Control del intercambio de sustancias: la membrana regula qué moléculas entran y salen de la célula, cuándo y en qué cantidad. Esta función es la base del mantenimiento de la homeostasis celular.
- Intercambio de información: la membrana recibe señales del entorno a través de receptores específicos y las traduce en respuestas intracelulares. Sin esta función, la coordinación entre células de un organismo multicelular sería imposible.
- Adhesión y reconocimiento celular: mediante moléculas de adhesión y el glicocáliz, la membrana permite que las células se reconozcan entre sí, se adhieran y se organicen en tejidos.
- Anclaje al citoesqueleto y a la matriz extracelular: la membrana conecta el interior de la célula con su entorno físico, transmitiendo fuerzas mecánicas y manteniendo la forma celular.
En este tema se desarrollan las dos primeras funciones: el intercambio de sustancias y el intercambio de información.
5.2. Transporte de sustancias a través de la membrana
El transporte de moléculas a través de la membrana puede ocurrir por dos vías: a través de la fase lipídica o a través de proteínas de transporte. En los procesos de permeabilidad simple no se producen cambios morfológicos visibles de la membrana; en la citosis (exo y endocitosis) sí.
5.2.1. Transporte a través de la fase lipídica
El interior hidrofóbico de la bicapa lipídica actúa como barrera para las moléculas polares y cargadas, pero permite el paso libre de las moléculas liposolubles.
Las moléculas que atraviesan la membrana por difusión simple a través de la fase lipídica son:
- Gases como O₂, CO₂ y N₂, que difunden libremente por ser apolares.
- Moléculas pequeñas polares sin carga, como el agua, la urea y el glicerol, que difunden con relativa facilidad pese a su polaridad.
- Moléculas liposolubles como el benceno, las hormonas esteroideas y la hormona tiroidea.
En cambio, las moléculas grandes polares sin carga (glucosa, sacarosa) y todas las moléculas cargadas (iones) no pueden atravesar la fase lipídica y necesitan proteínas de transporte específicas.
| Tipo de molécula | Ejemplos | Permeabilidad | Mecanismo de paso |
|---|---|---|---|
| Gases y moléculas apolares | O₂, CO₂, N₂, benceno, hormonas esteroideas | Muy alta | Difusión simple a través de la fase lipídica |
| Moléculas pequeñas polares sin carga | H₂O, urea, glicerol | Alta | Difusión simple · agua también por acuaporinas |
| Moléculas grandes polares sin carga | Glucosa, sacarosa, aminoácidos | Muy baja | Requieren proteínas transportadoras (permeasas) |
| Iones y moléculas cargadas | Na⁺, K⁺, Ca²⁺, Cl⁻, H⁺ | Prácticamente nula | Solo a través de canales iónicos o transportadores |
5.2.2. Transporte a través de proteínas: canales y transportadores
Las proteínas de membrana implicadas en el transporte se clasifican en dos grandes grupos:
- Canales iónicos: son proteínas transmembrana que forman un poro acuoso a través de la bicapa. Permiten el paso selectivo de iones específicos a favor de gradiente electroquímico, sin gasto energético directo. La selectividad del canal depende del diámetro del poro y de las cargas de los aminoácidos que lo revisten. Los canales pueden estar siempre abiertos o regularse por ligandos, voltaje o cambios mecánicos.
- Proteínas transportadoras o permeasas: son proteínas que se unen a la molécula a transportar y sufren un cambio de conformación que la traslada al otro lado de la membrana. Son más lentas que los canales pero más selectivas y pueden transportar moléculas más grandes.
5.2.3. Tipos de transporte: uniporte, simporte y antiporte
Las permeasas pueden transportar una sola molécula (uniporte) o acoplar el transporte de dos moléculas simultáneamente (cotransporte).
En el simporte, las dos moléculas se transportan en el mismo sentido. El ejemplo más conocido es el cotransportador Na⁺/glucosa del intestino: el Na⁺ entra a favor de su gradiente electroquímico y arrastra la glucosa hacia el interior celular en contra de su gradiente de concentración.
En el antiporte, las dos moléculas se transportan en sentidos opuestos. El intercambiador Na⁺/Ca²⁺ de la membrana plasmática es un ejemplo: la entrada de Na⁺ a favor de gradiente impulsa la salida de Ca²⁺ en contra de gradiente.
Este acoplamiento permite transportar sustancias en contra de su gradiente sin gastar ATP directamente, aprovechando el gradiente de otra molécula generado previamente por transporte activo primario (como la bomba Na⁺/K⁺ ATPasa).
Los mecanismos moleculares del transporte activo primario, los canales iónicos voltaje-dependientes y su papel en la generación del potencial de membrana se desarrollan en Fisiología General, concretamente en T4 — Membranas celulares y mecanismos de transporte y T9 — Propiedades de la membrana en reposo.
5.3. Intercambio de información: señalización celular
Además del intercambio de sustancias, la membrana plasmática es la plataforma principal de recepción de señales del entorno. La señalización celular permite a las células comunicarse, coordinarse y responder a cambios del medio.
5.3.1. Tipos de señalización según la distancia
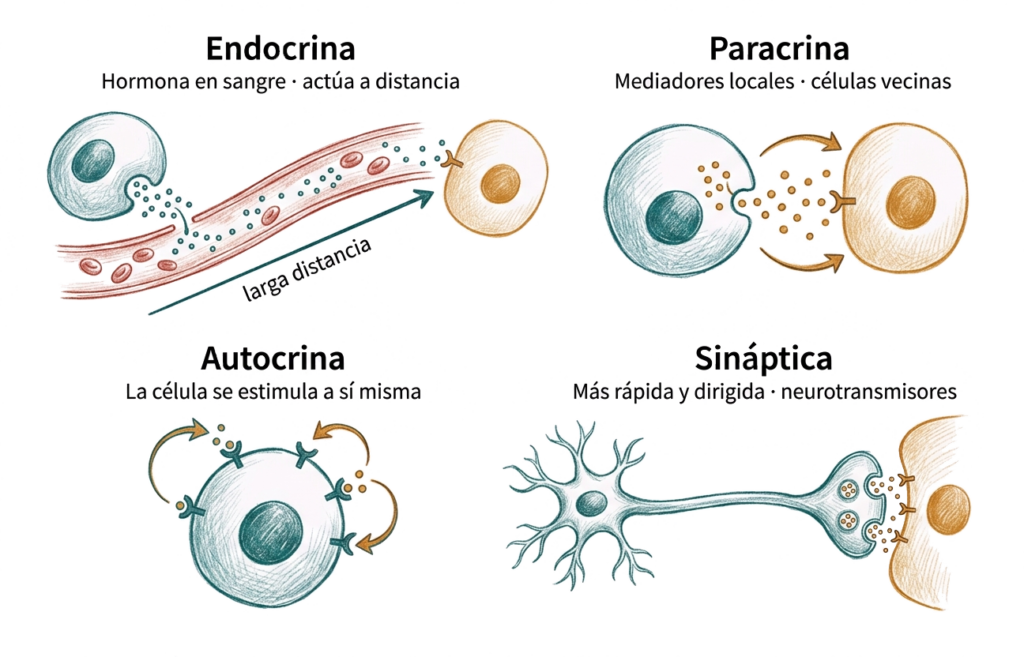
Según la distancia a la que actúan las moléculas señalizadoras y su vía de transporte, se distinguen cuatro tipos:
- Señalización endocrina: las células endocrinas secretan moléculas señalizadoras (hormonas) que se vierten al torrente circulatorio y actúan sobre células diana situadas a distancia. Es lenta pero tiene un alcance amplio.
- Señalización paracrina: las células secretan mediadores locales que actúan sobre células vecinas sin entrar en la sangre. Los mediadores tienen vida media corta, lo que limita su radio de acción. Las prostaglandinas y los factores de crecimiento locales son ejemplos.
- Señalización autocrina: la célula produce una molécula señalizadora para la que ella misma tiene receptor, estimulándose a sí misma. Es especialmente relevante durante el desarrollo embrionario para amplificar respuestas de diferenciación.
- Transmisión sináptica: es el tipo de señalización más rápido y más dirigido. La señal recorre el axón como una onda de despolarización y actúa localmente en la sinapsis mediante un mecanismo paracrino. Permite la comunicación a largas distancias (axones de hasta 1 metro) con alta precisión.
5.3.2. Tipos de receptores
Los receptores de las moléculas señalizadoras se clasifican según su localización:
- Receptores de superficie: son proteínas transmembrana que captan el ligando en el exterior celular y transmiten la señal al interior sin que el ligando atraviese la membrana. Sus ligandos son moléculas hidrofílicas que no pueden cruzar la bicapa: péptidos, proteínas, neurotransmisores y la mayoría de las hormonas no esteroideas.
- Receptores intracelulares: se encuentran en el citoplasma o en el núcleo. Sus ligandos son moléculas lipofílicas capaces de atravesar la bicapa por difusión simple: hormonas esteroideas (cortisol, estrógenos, testosterona), hormona tiroidea y vitamina D. El complejo ligando-receptor actúa generalmente como factor de transcripción, modificando directamente la expresión génica.
Los mecanismos moleculares de transducción de señales, las proteínas G y los segundos mensajeros se desarrollan en T7 de Biología — Principios básicos de señalización intracelular y en los temas de señalización de Bioquímica.
5.4. Exocitosis
La exocitosis es el proceso por el que una célula libera moléculas al exterior fusionando vesículas internas con la membrana plasmática. Es el mecanismo de secreción principal de proteínas, lípidos y otras macromoléculas.
El proceso general es: síntesis en el retículo endoplásmico rugoso → modificación y empaquetamiento en el aparato de Golgi → transporte en vesículas → fusión con la membrana plasmática → liberación del contenido al espacio extracelular.
Existen dos modalidades:
- Exocitosis constitutiva.
- Exocitosis regulada.
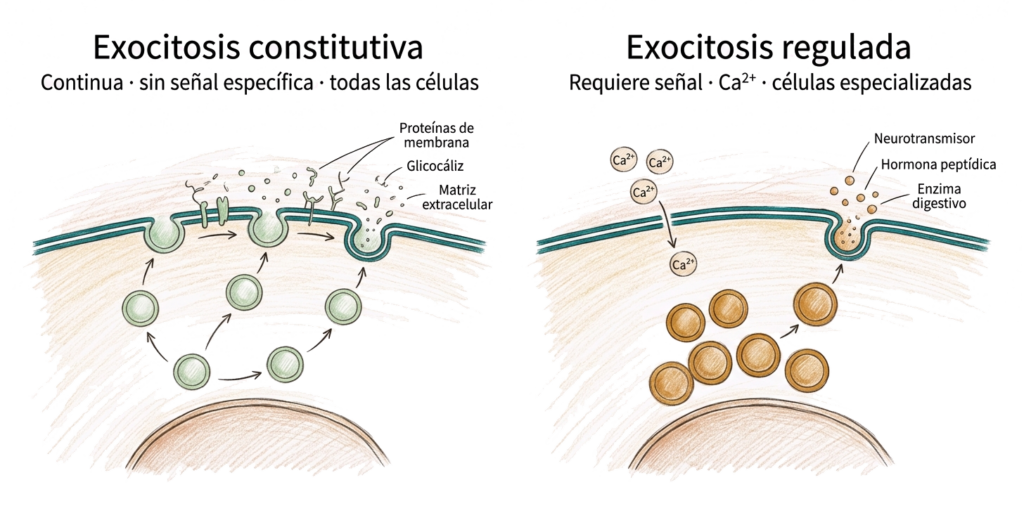
5.4.1. Exocitosis constitutiva
La exocitosis constitutiva ocurre de forma continua e independiente de señales externas. Las vesículas se fusionan con la membrana plasmática en cuanto llegan, sin necesidad de ningún estímulo. Es la vía por defecto de secreción: todas las células la realizan.
Sus productos incluyen componentes del glicocáliz (glucoproteínas y glicolípidos que se incorporan a la membrana), proteínas de la matriz extracelular (colágeno, fibronectina) y proteínas de señalización local.
5.4.2. Exocitosis regulada
La exocitosis regulada requiere una señal específica para desencadenarse. Las vesículas secretoras se acumulan en el citoplasma y solo se fusionan con la membrana cuando llega la señal adecuada, habitualmente un aumento de Ca²⁺ intracelular.
Es el mecanismo de secreción de las células especializadas: neuronas (liberación de neurotransmisores), células endocrinas (secreción de hormonas peptídicas), células exocrinas (liberación de enzimas digestivos) y mastocitos (liberación de histamina).
5.5. Endocitosis
La endocitosis es el proceso inverso a la exocitosis: la célula incorpora material del exterior formando vesículas a partir de invaginaciones de la membrana plasmática. A diferencia del transporte a través de proteínas, la endocitosis implica cambios morfológicos visibles de la membrana.
| Tipo | Material incorporado | Selectividad | Células que lo realizan | Vesícula resultante |
|---|---|---|---|---|
| Fagocitosis | Partículas sólidas grandes: bacterias, células muertas, restos celulares | Inespecífica (reconocimiento de patrones) | Macrófagos, neutrófilos, células dendríticas | Fagosoma (grande, >1 μm) |
| Pinocitosis | Líquido extracelular con solutos disueltos | Inespecífica | Prácticamente todas las células | Vesícula pequeña (<150 nm) |
| Endocitosis mediada por receptores | Ligandos específicos reconocidos por receptores de membrana | Alta — solo el ligando del receptor | Prácticamente todas las células | Vesícula recubierta de clatrina (~100 nm) |
| Transcitosis | Moléculas grandes (proteínas, anticuerpos) | Variable | Células endoteliales, epitelio intestinal neonatal | Vesícula de transcitosis |
5.5.1. Fagocitosis
La fagocitosis es la incorporación de partículas sólidas grandes: bacterias, células muertas, restos celulares. Solo la realizan células especializadas: macrófagos, neutrófilos y células dendríticas.
El proceso implica la extensión de prolongaciones citoplasmáticas (pseudópodos) que rodean la partícula y la engullen formando una vesícula grande denominada fagosoma. El fagosoma se fusiona con lisosomas, cuyas enzimas degradan el contenido.
5.5.2. Pinocitosis
La pinocitosis es la incorporación de pequeñas gotas de líquido extracelular con los solutos que contiene. La realizan prácticamente todas las células. Las vesículas formadas son mucho más pequeñas que los fagosomas.
Es un proceso inespecífico: la célula incorpora todo lo que está disuelto en el líquido extracelular próximo a la membrana, sin selección de moléculas concretas.
5.5.3. Endocitosis mediada por receptores
La endocitosis mediada por receptores es el mecanismo más selectivo y eficiente de internalización de moléculas específicas. Permite a la célula concentrar en el interior moléculas presentes a baja concentración en el líquido extracelular.
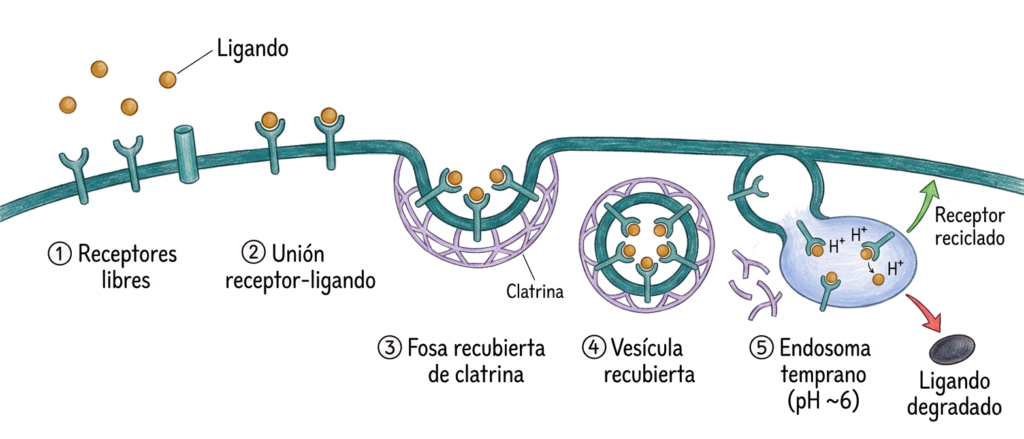
El proceso ocurre en fosas especializadas de la membrana plasmática denominadas fosas recubiertas de clatrina (coated pits):
Los receptores específicos de membrana reconocen y se unen a sus ligandos en el exterior celular. Los complejos receptor-ligando se concentran en las fosas recubiertas, donde la proteína clatrina forma una red en la cara citosólica que deforma la membrana hacia el interior. La invaginación se cierra formando una vesícula recubierta de clatrina que se desprende de la membrana.
La clatrina se disocia rápidamente de la vesícula, que se fusiona con el endosoma temprano. En el endosoma, el pH desciende a ~6 por acción de bombas de H⁺. Este descenso del pH provoca la disociación del ligando y el receptor.
Desde el endosoma, los destinos posibles son dos: el receptor puede reciclarse de vuelta a la membrana plasmática (para ser usado de nuevo) o degradarse en los lisosomas. El ligando puede ser degradado en lisosomas, reciclado o transcitosado.
La hipercolesterolemia familiar es el ejemplo clásico de enfermedad por defecto en la endocitosis mediada por receptores. Las células normales internalizan el colesterol unido a LDL mediante el receptor de LDL, que se recicla tras liberar la LDL en el endosoma. Las mutaciones en el receptor de LDL impiden la internalización de LDL, que se acumula en sangre produciendo niveles de colesterol muy elevados y enfermedad cardiovascular precoz. Fue el modelo que permitió a Brown y Goldstein describir el mecanismo de la endocitosis mediada por receptores, trabajo por el que recibieron el Nobel de Fisiología en 1985.
5.5.4. Transcitosis
La transcitosis es un proceso especial en el que una célula internaliza una molécula por endocitosis en un lado y la libera por exocitosis en el lado opuesto, transportando la molécula a través de la célula sin degradarla.
Es especialmente relevante en las células endoteliales de los capilares, donde permite el transporte de proteínas grandes desde la sangre al tejido intersticial, y en las células epiteliales intestinales, donde permite el paso de anticuerpos maternos (IgA) desde la luz intestinal a la circulación en el recién nacido.
