12.1. Funciones generales del citoesqueleto
El citoesqueleto integra cuatro funciones principales en la célula eucariota:
- Control de la posición de estructuras intracelulares: los orgánulos no flotan libremente en el citoplasma sino que están anclados y posicionados por los filamentos del citoesqueleto. El RE se extiende a lo largo de los microtúbulos hacia la periferia; el aparato de Golgi se mantiene cerca del centrosoma gracias a la dineína citoplasmática; las mitocondrias se localizan donde más se necesita ATP.
- Control de la forma celular: el córtex de actina y los filamentos intermedios determinan la forma celular y su resistencia mecánica. Las células cambian de forma mediante reorganizaciones controladas del citoesqueleto de actina.
- Movimiento celular e intracelular: el transporte de vesículas, orgánulos y ARNm a lo largo de microtúbulos y microfilamentos, y la migración celular sobre sustratos, dependen del citoesqueleto y sus proteínas motoras.
- División celular: el huso mitótico (microtúbulos) segrega los cromosomas; el anillo contráctil (actina y miosina) divide la célula en dos.
12.2. Proteínas motoras
Las proteínas motoras son proteínas que se unen a filamentos del citoesqueleto y utilizan la energía de la hidrólisis de ATP para generar movimiento unidireccional. Tienen tres dominios funcionales: la cabeza motora (se une al filamento y contiene la actividad ATPasa), el cuello (transmite el cambio conformacional) y la cola (se une a la carga: vesícula, orgánulo u otro filamento).
12.2.1. Kinesinas: transporte anterógrado en microtúbulos
Las kinesinas son proteínas motoras que se desplazan hacia el extremo (+) de los microtúbulos, es decir, desde el centrosoma hacia la periferia celular (transporte anterógrado).
La kinesina convencional es un tetrámero formado por dos cadenas pesadas (con dos cabezas globulares que son los dominios motores) y dos cadenas ligeras (que se unen a la carga). Las dos cabezas actúan de forma coordinada y alternante, como «pasos» a lo largo del microtúbulo, avanzando ~8 nm por ciclo de hidrólisis de ATP.
Las kinesinas transportan vesículas del aparato de Golgi hacia la membrana plasmática, orgánulos hacia la periferia y componentes sinápticos a lo largo de los axones en el transporte axonal anterógrado.
En mamíferos existen más de 40 kinesinas distintas con especificidades de carga diferentes.
12.2.2. Dineínas: transporte retrógrado en microtúbulos
Las dineínas se desplazan hacia el extremo (-) de los microtúbulos, es decir, desde la periferia hacia el centrosoma (transporte retrógrado). Son los motores moleculares más grandes conocidos.
Existen dos tipos principales:
- Dineína citoplasmática: presente en prácticamente todas las células eucariotas. Media el transporte retrógrado de vesículas endosomales y lisosomales, el posicionamiento del aparato de Golgi cerca del centrosoma y el movimiento del núcleo durante la migración celular. Requiere el cofactor dinactina para unirse eficientemente a la carga.
- Dineína axonemal: especializada en el movimiento de cilios y flagelos. Sus brazos externos e internos generan el deslizamiento de los dobletes del axonema.
12.2.3. Miosinas: motores sobre microfilamentos
Las miosinas son proteínas motoras que se desplazan sobre los filamentos de actina. Existen más de 35 clases de miosinas en eucariotas; las más relevantes son:
- Miosina II: la miosina clásica del músculo y de las células no musculares. Es un hexámero formado por dos cadenas pesadas (con dos cabezas motoras y una cola helicoidal que permite la dimerización) y cuatro cadenas ligeras. Las moléculas de miosina II se ensamblan en filamentos bipolares que pueden generar fuerza contráctil sobre los filamentos de actina.
- Miosina V: miosina de doble cabeza con gran paso (~36 nm por ciclo). Transporta vesículas, orgánulos y ARNm a lo largo de los microfilamentos.
- Miosina I: monómero con un dominio de unión a lípidos. Ancla los filamentos de actina a la membrana plasmática en las microvellosidades.
| Proteína | Filamento | Dirección | Velocidad | Función principal | Ejemplo fisiológico |
|---|---|---|---|---|---|
| Kinesina convencional (KIF5) | Microtúbulos | Hacia extremo (+) · periferia · anterógrado | ~1 μm/s | Transporte de vesículas y orgánulos hacia la periferia | Transporte axonal anterógrado · distribución del Golgi |
| Dineína citoplasmática | Microtúbulos | Hacia extremo (-) · centrosoma · retrógrado | ~1-2 μm/s | Transporte retrógrado · posicionamiento del Golgi · movimiento nuclear | Transporte axonal retrógrado · posicionamiento del núcleo |
| Dineína axonemal | Microtúbulos del axonema | Hacia extremo (-) del doblete vecino | ~14 μm/s (in vitro) | Genera el movimiento ciliar y flagelar por deslizamiento de dobletes | Movimiento del espermatozoide · aclaramiento mucociliar |
| Miosina II | Microfilamentos | Hacia extremo (+) | Variable | Contracción muscular · citocinesis · fibras de estrés | Contracción del sarcómero · anillo contráctil |
| Miosina V | Microfilamentos | Hacia extremo (+) | ~0,5 μm/s | Transporte de vesículas y orgánulos a lo largo de actina | Transporte de melanosomas · vesículas sinápticas |
| Miosina I | Microfilamentos | Hacia extremo (+) | Baja | Ancla actina a membrana plasmática | Estructura de microvellosidades |
El ciclo mecano-químico de la miosina II tiene cuatro pasos: unión al filamento de actina sin nucleótido (estado de rigor), unión de ATP que reduce la afinidad por la actina, hidrólisis de ATP a ADP+Pi que produce el cambio conformacional prepotencia, y unión débil a un nuevo sitio de actina seguida de liberación de Pi que genera el golpe de potencia (~5 nm de desplazamiento) y libera ADP.
12.3. Movimiento celular
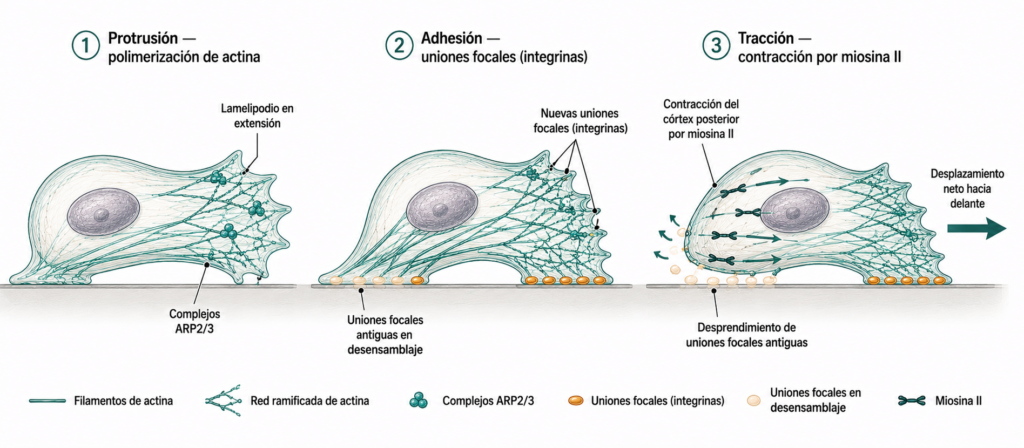
La migración celular sobre superficies sólidas es un proceso complejo que implica la coordinación de tres actividades:
- Protrusión: en el frente de avance, la polimerización de actina impulsada por el complejo ARP2/3 genera lamelipodios que se extienden hacia adelante. La presión osmótica local y la polimerización de actina empujan la membrana.
- Adhesión: el lamelipodio forma nuevas uniones focales mediante integrinas que anclan la célula al sustrato en el frente de avance. Sin adhesión, la protrusión no genera movimiento neto.
- Tracción: la contracción del córtex de actina posterior por la miosina II desplaza el cuerpo celular hacia adelante y desensambla las adhesiones antiguas en la parte posterior. La célula avanza tirando de sí misma hacia el frente.
Este ciclo de protrusión-adhesión-tracción se repite continuamente y puede ocurrir de forma muy coordinada (células epiteliales deslizándose suavemente) o de forma más irregular (fibroblastos con movimiento en pasos).
12.4. Regulación del citoesqueleto por las GTPasas Rho
La reorganización del citoesqueleto de actina en respuesta a señales extracelulares está coordinada por las GTPasas de la familia Rho, pequeñas proteínas G monoméricas que alternan entre una forma activa unida a GTP y una forma inactiva unida a GDP.
Las tres GTPasas principales tienen efectos distintos y bien definidos sobre el citoesqueleto de actina:
- RhoA: activa cuando la célula recibe señales de adhesión a la matriz extracelular o de factores de crecimiento. Promueve la formación de fibras de estrés (haces contráctiles de actina y miosina II) y de uniones focales (complejos de adhesión a la matriz). Hace que la célula se «tense» y adhiera firmemente al sustrato.
- Rac1: activa en el frente de avance de las células en migración. Activa el complejo ARP2/3 y promueve la formación de lamelipodios y de uniones adherentes con células vecinas.
- Cdc42: activa en respuesta a gradientes de quimiocinas y factores de crecimiento. Activa la nucleación de filamentos de actina paralelos y promueve la formación de filopodios, extensiones digitiformes que exploran el entorno y detectan gradientes químicos.
Las tres GTPasas no actúan de forma independiente: Cdc42 puede activar Rac, que a su vez puede activar Rho, formando una cascada de señalización que coordina la reorganización espacial del citoesqueleto.
| GTPasa | Estructura de actina activada | Señal activadora | Función fisiológica principal |
|---|---|---|---|
| RhoA | Fibras de estrés · uniones focales | Adhesión a matriz extracelular · lisofosfolípidos (LPA) | Tensión celular · adhesión firme al sustrato · citocinesis |
| Rac1 | Lamelipodios · uniones adherentes | Factores de crecimiento · integrinas · Cdc42 (cascada) | Migración celular · extensión del frente de avance · formación de epitelio |
| Cdc42 | Filopodios | Gradientes de quimiocinas · factores de crecimiento locales | Exploración del entorno · guía axonal · polaridad celular |
12.5. El citoesqueleto en la división celular
12.5.1. El huso mitótico
Durante la mitosis, los microtúbulos citoplasmáticos se desensamblan y se reorganizan formando el huso mitótico, una estructura bipolar con dos polos (centrosomas) y tres tipos de microtúbulos:
- Microtúbulos astrales: irradian desde cada centrosoma hacia la corteza celular y posicionan el huso en el centro de la célula.
- Microtúbulos polares: se solapan en la zona ecuatorial con los del polo opuesto y son empujados entre sí por kinesinas bipolares, separando los polos del huso y elongando la célula.
- Microtúbulos cinetocóricos: se unen a los cinetocoros de los cromosomas (uno por cada cromátida hermana) y, mediante la depolimerización del extremo (+) y la acción de la dineína, arrastran los cromosomas hacia los polos.
12.5.2. El anillo contráctil y la citocinesis
Tras la segregación de los cromosomas, la célula se divide en dos por citocinesis. En el ecuador celular se ensambla el anillo contráctil: una banda de filamentos de actina y miosina II dispuestos perpendicularmente al eje del huso. La posición del anillo está determinada por señales del huso mitótico.
La miosina II contrae el anillo de actina progresivamente, estrangulando la célula hasta dividirla en dos células hijas. La membrana plasmática sigue el anillo hacia el interior, formando el surco de división, hasta que las dos células quedan unidas solo por el cuerpo medio, un puente citoplasmático que finalmente se rompe completando la citocinesis.
