19.1. La meiosis y la reproducción sexual
La reproducción sexual permite mezclar el material genético de dos individuos distintos, generando descendientes que difieren entre sí y de sus progenitores. Esta variabilidad es la base de la adaptación evolutiva de las especies.
A nivel celular, la reproducción sexual exige una alternancia entre células diploides (2n) y células haploides (n). Las células haploides, llamadas gametos, se fusionan en la fecundación para formar el cigoto diploide, que mediante mitosis originará el nuevo organismo. Para que el número de cromosomas no se duplique en cada generación, los gametos deben formarse por un mecanismo especial que reduzca la dotación cromosómica a la mitad: la meiosis.
Meiosis: proceso de división celular que genera cuatro células haploides (n) genéticamente distintas a partir de una célula diploide (2n), mediante dos divisiones sucesivas sin replicación intermedia del ADN.
19.2. Visión general de la meiosis
La meiosis consta de dos divisiones sucesivas denominadas meiosis I y meiosis II, separadas por una interfase muy breve en la que no se replica el ADN.
- La meiosis I es la división reduccional: los cromosomas homólogos se separan, reduciendo la dotación de 2n a n. Las células resultantes son haploides pero contienen cromosomas con dos cromátidas.
- La meiosis II es la división ecuacional: equivale funcionalmente a una mitosis. Las cromátidas hermanas de cada cromosoma se separan, generando cuatro células haploides con cromosomas de una sola cromátida.
Lo que distingue la meiosis de dos mitosis consecutivas está en la profase I: el apareamiento de cromosomas homólogos, la recombinación y la formación de quiasmas son eventos exclusivos de la meiosis que no ocurren en ninguna fase de la mitosis.
19.3. Meiosis I: la división reduccional
19.3.1. Profase I
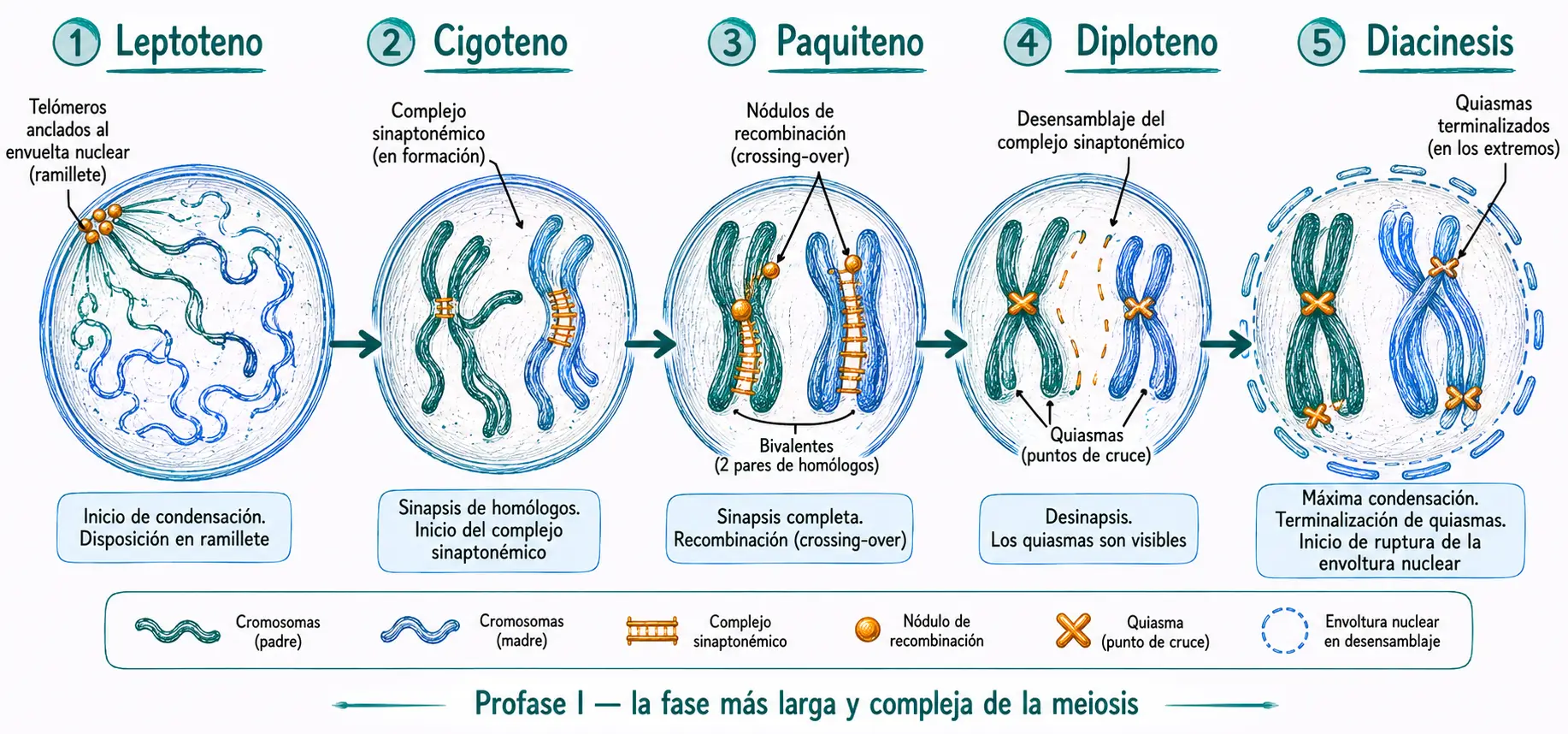
La profase I es la fase más larga y compleja de toda la meiosis. Puede durar desde horas hasta años. En los ovocitos humanos, las células permanecen detenidas en diploteno desde la vida fetal hasta la ovulación. Se divide en cinco subfases: leptoteno, cigoteno, paquiteno, diploteno y diacinesis.
Leptoteno
Los cromosomas comienzan a condensarse y se individualizan. Aunque ya tienen dos cromátidas (la replicación ocurrió en la fase S previa), solo se visualiza una por el grado de condensación. Los telómeros se anclan a la envoltura nuclear a través de estructuras proteicas llamadas placas de adhesión, adoptando los cromosomas una disposición característica en «ramillete» o bouquet. El nucléolo permanece intacto. Aparecen los primeros nódulos de recombinación, complejos proteicos multienzimáticos precursores del complejo sinaptonémico, de los que pueden existir hasta 500 en esta fase.
Cigoteno
Cada cromosoma comienza a reconocer y alinearse con su homólogo a lo largo de toda su longitud. El cromosoma paterno busca al materno y los dos se aproximan de forma progresiva, como el cierre de una cremallera. El resultado es la formación de los bivalentes (también llamados tétradas): estructuras formadas por los dos cromosomas homólogos con sus respectivas cromátidas hermanas, es decir, cuatro cromátidas en total.
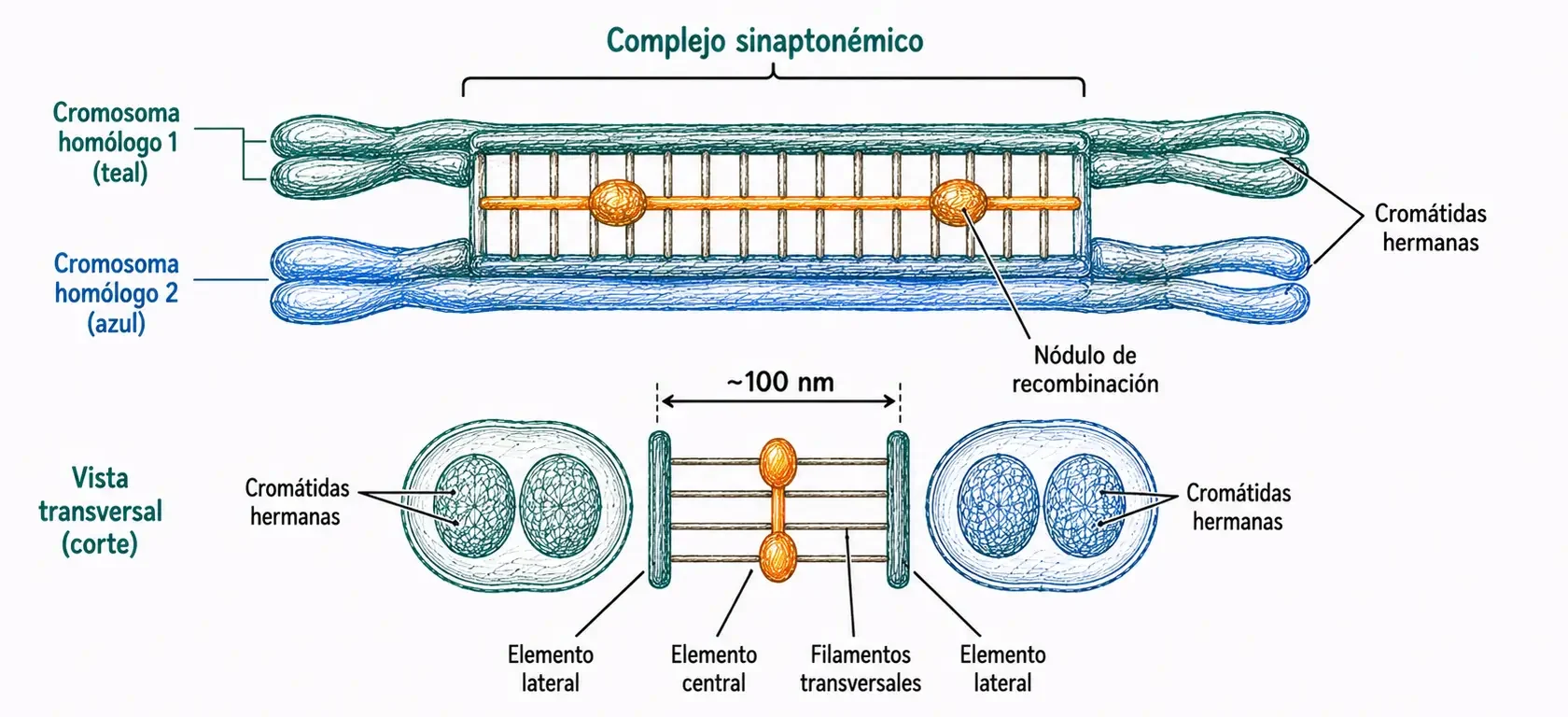
Este apareamiento preciso entre homólogos se facilita por la formación del complejo sinaptonémico, una estructura proteica con forma de escalera que se ensambla entre los dos homólogos a medida que se aparean.
Los cromosomas sexuales X e Y, que no son totalmente homólogos, se aparean únicamente en sus regiones pseudoautosómicas: una mayor en el extremo del brazo corto (~2,4 Mb) y una menor en el extremo del brazo largo (~300 kb). La recombinación entre X e Y ocurre casi exclusivamente en la región mayor.
Paquiteno
El apareamiento entre homólogos se completa a lo largo de toda la longitud cromosómica. El complejo sinaptonémico queda totalmente ensamblado, formando una estructura trilateral: dos elementos laterales (uno por cada par de cromátidas hermanas) conectados por un elemento central mediante filamentos transversales. El diámetro total del complejo es de ~100 nm.
En esta fase se produce la recombinación genética (también llamada crossing-over o entrecruzamiento): se introducen roturas de doble cadena en el ADN, que en lugar de repararse localmente se resuelven mediante intercambio de segmentos entre cromátidas no hermanas de los dos homólogos. Este proceso está catalizado por los nódulos de recombinación, que en paquiteno ya han disminuido a unos 20 por bivalente.
El complejo sinaptonémico no cataliza directamente la recombinación, pero la dirige de forma que ocurra entre cromátidas de homólogos distintos y no entre cromátidas hermanas del mismo cromosoma. Sin complejo sinaptonémico no hay recombinación meiótica eficiente.
Diploteno
El complejo sinaptonémico se desensambla. Los cromosomas homólogos se separan parcialmente, pero permanecen unidos en los puntos donde se produjo la recombinación: los quiasmas. Un quiasma es la huella morfológica visible del entrecruzamiento entre cromátidas homólogas.
El número de quiasmas por bivalente varía según la especie y la longitud del cromosoma, pero siempre existe al menos uno por par de homólogos. Este mínimo no es arbitrario:
Sin al menos un quiasma por bivalente, los homólogos no pueden segregar correctamente en anafase I, lo que genera aneuploidías. Los quiasmas actúan como puntos de tensión que permiten al huso detectar y mantener la alineación bipolar de cada bivalente.
La formación de un quiasma reduce la probabilidad de que se forme otro en regiones próximas del mismo cromosoma, fenómeno denominado interferencia. Este mecanismo garantiza que incluso los cromosomas cortos formen al menos un quiasma.
En diploteno, la cromatina no está totalmente condensada y el ADN puede transcribirse activamente. Es en esta fase donde los cromosomas plumosos de los ovocitos de anfibios (descritos en el Tema 15 – El cromosoma metafásico) alcanzan su máxima actividad transcripcional.
Algunos ovocitos humanos permanecen detenidos en diploteno durante décadas. Esta detención prolongada explica en parte el aumento del riesgo de errores de segregación con la edad materna.
Diploteno
La cromatina se condensa más, y las dos cromátidas de cada homólogo son ya visibles por separado. Los quiasmas se desplazan progresivamente desde las regiones internas hacia los telómeros de los cromosomas: este proceso se denomina terminalización y continúa en la prometafase I.
19.3.2. Prometafase I
La envoltura nuclear se rompe y los cromosomas son accesibles a los microtúbulos del huso. Los quiasmas terminalizan completamente. La diferencia respecto a la prometafase mitótica es que los cromosomas se presentan en bivalentes: cada bivalente debe quedar unido a microtúbulos de ambos polos para alinearse en el ecuador.
A diferencia de la mitosis, los cinetocoros de las cromátidas hermanas de un mismo homólogo funcionan como una unidad y se orientan hacia el mismo polo. Los microtúbulos de un polo capturan el bivalente por un lado; los del polo opuesto lo capturan por el otro, estableciendo la tensión necesaria para la alineación ecuatorial.
19.3.3. Metafase I
Los bivalentes se alinean en la placa metafásica. La orientación de cada bivalente en el plano ecuatorial es completamente al azar: el cromosoma paterno puede quedar hacia cualquiera de los dos polos, independientemente de cómo se orienten los otros pares. Esta independencia es la base de la segregación aleatoria.
| Característica | Metafase I (meiosis) | Metafase (mitosis) |
|---|---|---|
| Unidad en la placa | Bivalente (4 cromátidas) | Cromosoma individual (2 cromátidas) |
| Orientación de cinetocoros hermanos | Hacia el mismo polo | Hacia polos opuestos |
| Qué se separa en anafase | Cromosomas homólogos | Cromátidas hermanas |
19.3.4. Anafase I
La separación de los homólogos se desencadena por la degradación de las cohesinas de los brazos cromosómicos por la separasa, lo que permite que los quiasmas se resuelvan y los homólogos se alejen hacia polos opuestos.
Las cohesinas centroméricas, sin embargo, no se degradan en anafase I: permanecen intactas y mantienen unidas las dos cromátidas de cada homólogo. Serán escindidas en la anafase II.
La segregación de cada par de homólogos hacia un polo u otro es independiente del resto de pares. Con 23 pares cromosómicos humanos, esto genera 2²³ (~8 millones) combinaciones posibles de cromosomas en cada gameto, solo por segregación aleatoria.
19.3.5. Telofase I e interfase
Los dos lotes de cromosomas haploides (cada uno con cromosomas de dos cromátidas) llegan a los polos. Se produce la citocinesis, formando dos células haploides. En muchas especies se reconstituyen brevemente las envolturas nucleares y el nucléolo.
La interfase entre meiosis I y meiosis II es muy breve y, crucialmente, no hay replicación del ADN: las células pasan directamente a la profase II.
19.4. Meiosis II: la división ecuacional
La meiosis II es funcionalmente equivalente a una mitosis, pero parte de células haploides con cromosomas de dos cromátidas recombinadas.
- En la profase II, los cromosomas se recondensan si se habían descondensado.
- En la prometafase II, la envoltura nuclear se fragmenta y los cromosomas son capturados por el huso. En esta división, los cinetocoros de las cromátidas hermanas actúan de forma independiente y se orientan hacia polos opuestos, exactamente igual que en la mitosis.
- En la metafase II, los cromosomas se alinean individualmente en la placa ecuatorial.
- En la anafase II, las cohesinas centroméricas son escindidas por la separasa y las cromátidas hermanas se separan hacia los polos.
- En la telofase II, se reconstituyen las envolturas nucleares y se produce la citocinesis.
El resultado final son cuatro células haploides (n), cada una con cromosomas de una sola cromátida, genéticamente distintas entre sí.
19.5. Variabilidad genética generada por la meiosis
La meiosis es la principal fuente de variabilidad genética en los organismos de reproducción sexual. Esta variabilidad surge de dos mecanismos independientes y aditivos:
- Segregación aleatoria de homólogos. En metafase I, la orientación de cada bivalente hacia uno u otro polo es independiente del resto. Con 23 pares cromosómicos, esto genera 2²³ combinaciones posibles de cromosomas paternos y maternos en cada gameto, incluso sin recombinación.
- Recombinación genética. Durante el paquiteno, el intercambio de segmentos entre cromátidas no hermanas crea cromosomas recombinados que contienen fragmentos de ambos progenitores. Cuantos más quiasmas se formen (a mayor longitud cromosómica), mayor es la variabilidad generada.
Además, en la fecundación, cada gameto se une con otro que aporta su propio conjunto de combinaciones, multiplicando exponencialmente la variabilidad del nuevo individuo.
19.6. Evolución del material genético durante la meiosis
| Fase | Ploidía | Cromátidas/cromosoma | Cantidad ADN |
|---|---|---|---|
| G1 (premeiosis) | 2n | 1 | 2C |
| S / G2 (premeiosis) | 2n | 2 | 4C |
| Profase I – Metafase I | 2n | 2 | 4C |
| Anafase I | 2n* | 2 | 4C |
| Telofase I / células post-meiosis I | n | 2 | 2C |
| Profase II – Metafase II | n | 2 | 2C |
| Anafase II | 2n* | 1 | 2C |
| Telofase II / células hijas | n | 1 | C |
Los valores de ploidía en anafase I y II son transitorios: antes de completarse la citocinesis, la célula contiene cromosomas que aún no se han distribuido en núcleos separados. Al finalizar cada citocinesis se recupera la ploidía correcta.
19.7. Comparación entre mitosis y meiosis
| Característica | Mitosis | Meiosis |
|---|---|---|
| Número de divisiones | 1 | 2 |
| Células resultantes | 2 diploides (2n) | 4 haploides (n) |
| Identidad genética | Idénticas a la célula madre | Genéticamente distintas |
| Apareamiento de homólogos | No | Sí (profase I) |
| Recombinación | No | Sí (paquiteno) |
| Duración | 30 min – 2 h | Días a años |
| Función | Crecimiento, renovación tisular | Formación de gametos |
19.8. Errores meióticos y consecuencias clínicas
Aneuploidías por no disyunción
El error más frecuente de la meiosis es la no disyunción: el fallo en la separación de homólogos (meiosis I) o de cromátidas hermanas (meiosis II) en anafase. El resultado son gametos con un cromosoma de más (n+1) o de menos (n-1). Si estos gametos se fecundan, el cigoto tendrá 47 o 45 cromosomas en lugar de 46.
Las consecuencias más conocidas son las trisomías: la trisomía 21 (síndrome de Down), la trisomía 18 (síndrome de Edwards) y la trisomía 13 (síndrome de Patau). La mayoría de las monosomías autosómicas son letales en estadios precoces del desarrollo. Las aneuploidías de los cromosomas sexuales (síndrome de Turner 45,X; síndrome de Klinefelter 47,XXY) son más frecuentes y compatibles con la vida.
El riesgo de no disyunción aumenta con la edad materna, probablemente por el deterioro de las cohesinas y del complejo sinaptonémico durante el prolongado arresto en diploteno.
Recombinación desigual
Cuando el entrecruzamiento ocurre entre regiones no homólogas de cromosomas paralogos (especialmente en regiones con secuencias repetidas), el resultado es una recombinación desigual: un cromosoma gana material y el otro lo pierde. Este mecanismo es responsable de deleciones y duplicaciones cromosómicas como la deleción 22q11 (síndrome de DiGeorge) o las duplicaciones en la región 17p12 responsables de la enfermedad de Charcot-Marie-Tooth tipo 1A.
