6.1. Características generales y ultraestructura
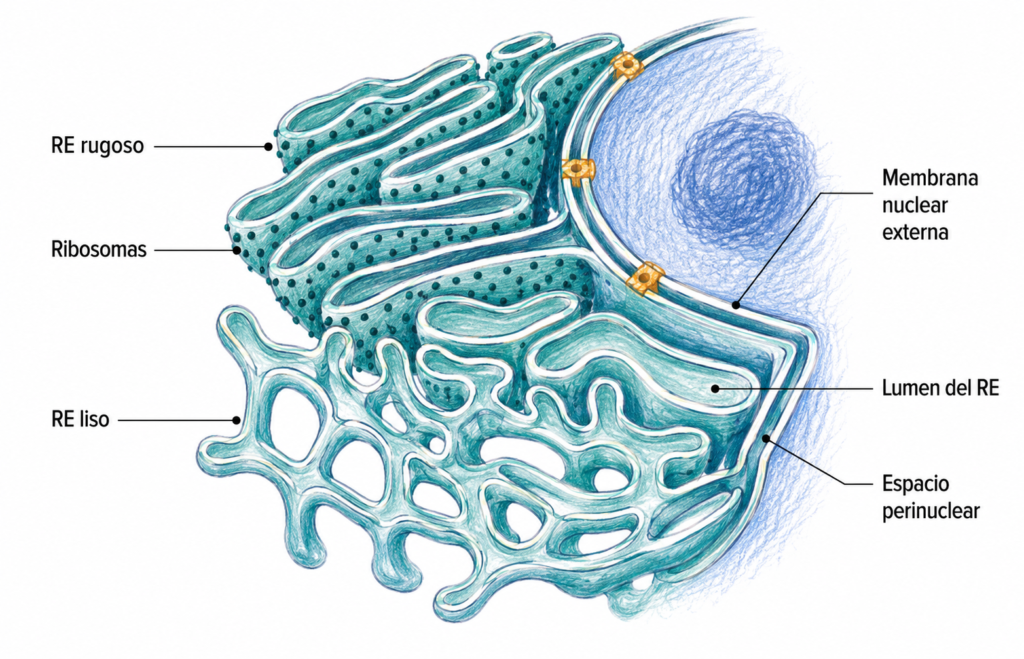
El retículo endoplásmico (RE) es el orgánulo membranoso más extenso de la célula eucariota. Forma una red continua de sáculos aplanados, cisternas y túbulos interconectados que ocupa gran parte del citoplasma y se conecta directamente con la membrana nuclear externa, de la que es continuación.
Al microscopio óptico, el RE no es visible como estructura individual, pero su presencia se manifiesta indirectamente: las zonas del citoplasma ricas en RE rugoso tienen afinidad por colorantes básicos (basofilia) debido a la alta concentración de ARN ribosomal. Al microscopio electrónico de transmisión, el RE aparece como una red de sáculos aplanados y túbulos delimitados por una membrana simple que encierra un espacio interno denominado lumen o espacio cisternal.
El RE establece dos compartimentos dentro de la célula: el citoplasma en el exterior de sus membranas y el lumen en su interior. El lumen del RE es continuo con el espacio perinuclear (entre las dos membranas nucleares), lo que permite el intercambio de contenido entre ambos compartimentos.
La membrana del RE tiene una composición similar a la membrana plasmática, con un 70% de proteínas y un 30% de lípidos, aunque prácticamente sin glúcidos. Las proteínas específicas de cada tipo de RE son las que confieren sus funciones características.
| Característica | RE rugoso | RE liso |
|---|---|---|
| Morfología | Cisternas y sáculos aplanados | Red de túbulos ramificados |
| Ribosomas | Sí — en cara citosólica | No |
| Función principal | Síntesis y procesamiento de proteínas | Síntesis de lípidos · detoxificación · almacén de Ca²⁺ |
| Proteínas sintetizadas | Proteínas de la vía secretora · proteínas transmembrana | No sintetiza proteínas |
| Abundante en | Células secretoras: páncreas exocrino · hepatocitos · células plasmáticas | Células esteroidogénicas · hepatocitos · músculo (REL = RS) |
| Relación con el núcleo | Continuo con la membrana nuclear externa | Continuo con el RE rugoso |
6.2. Retículo endoplásmico rugoso y liso
El RE no es homogéneo en toda su extensión. Se distinguen dos regiones morfológica y funcionalmente distintas, aunque continuas entre sí:
El RE rugoso (RER) recibe su nombre del aspecto granuloso que le confieren los ribosomas adheridos a su cara citosólica. Está formado principalmente por cisternas y sáculos aplanados. Es especialmente abundante en células con alta actividad secretora: células pancreáticas acinares, hepatocitos, fibroblastos y células plasmáticas. En estas células, el RER puede constituir hasta el 60% del área total de membrana celular.
El RE liso (REL) carece de ribosomas en su superficie y tiene predominantemente forma de red de túbulos ramificados. Es especialmente abundante en células especializadas en la síntesis de hormonas esteroideas (células de la corteza suprarrenal, células de Leydig del testículo) y en el metabolismo de lípidos. En el hígado, el REL participa en la detoxificación de fármacos y sustancias tóxicas.
Entre ambos existe el elemento transicional: una zona del RE con ribosomas solo en una cara, desde donde parten las vesículas que transportan proteínas hacia el aparato de Golgi.
La cantidad de RE presente en una célula varía con su estado metabólico. En células hepáticas de rata tratadas con fenobarbital u otros barbitúricos, el volumen del REL aumenta enormemente como respuesta adaptativa para metabolizar el fármaco. Al retirar el tratamiento, el exceso de REL se elimina por autofagia.
| Característica | N-glicosilación | O-glicosilación |
|---|---|---|
| Localización | Se inicia en el RE · se completa en el Golgi | Exclusivamente en el Golgi |
| Aminoácido receptor | Asparagina (Asn) en secuencia Asn-X-Ser/Thr | Serina (Ser) o Treonina (Thr) |
| Oligosacárido inicial | 14 residuos (2 GlcNAc + 9 Man + 3 Glc) transferidos desde dolicol | Un solo azúcar añadido secuencialmente |
| Función | Control de calidad del plegamiento · señal de dirección al lisosoma (M6P) | Protección de mucinas · lubricación · señalización |
| Ejemplos | Mayoría de glucoproteínas secretadas · enzimas lisosomales | Mucinas · proteoglicanos · O-GlcNAc en proteínas nucleares |
6.3. Funciones del RE rugoso
6.3.1. Síntesis de proteínas: el péptido señal y la SRP
El RER sintetiza todas las proteínas destinadas a la vía secretora: proteínas de secreción, proteínas de membrana de orgánulos y proteínas transmembrana de la membrana plasmática. También sintetiza los lípidos de membrana de mitocondrias y peroxisomas, aunque no sus proteínas.
La síntesis comienza siempre en ribosomas libres del citoplasma. Si la proteína está destinada al RE, los primeros aminoácidos sintetizados forman el péptido señal: una secuencia N-terminal de 16-30 aminoácidos predominantemente hidrofóbicos que actúa como señal de dirección al RE.
El péptido señal es reconocido por la partícula de reconocimiento de la señal (SRP), un complejo ribonucleoproteico citosólico. La SRP se une al péptido señal y detiene temporalmente la traducción, evitando que la cadena polipeptídica se libere al citoplasma. El complejo ribosoma-ARNm-cadena naciente-SRP se desplaza entonces hasta la membrana del RER, donde la SRP se une a su receptor (receptor de la SRP) en la membrana del RER.
Una vez anclado el ribosoma a la membrana, la SRP se disocia y la traducción se reanuda. La cadena polipeptídica en crecimiento se inserta en el complejo translocador SEC61, un canal acuoso formado por tres proteínas transmembrana que atraviesa la membrana del RER. La proteína se sintetiza directamente dentro del lumen del RE a medida que se alarga: es una síntesis cotraduccional.
Poco después de iniciarse la translocación, una peptidasa de señal en el lumen del RER corta y elimina el péptido señal. La proteína madura termina de sintetizarse en el lumen.
El destino final de la proteína depende de su estructura:
1. Las proteínas solubles (de secreción o de lumen de orgánulos) atraviesan completamente la membrana y quedan libres en el lumen del RE.
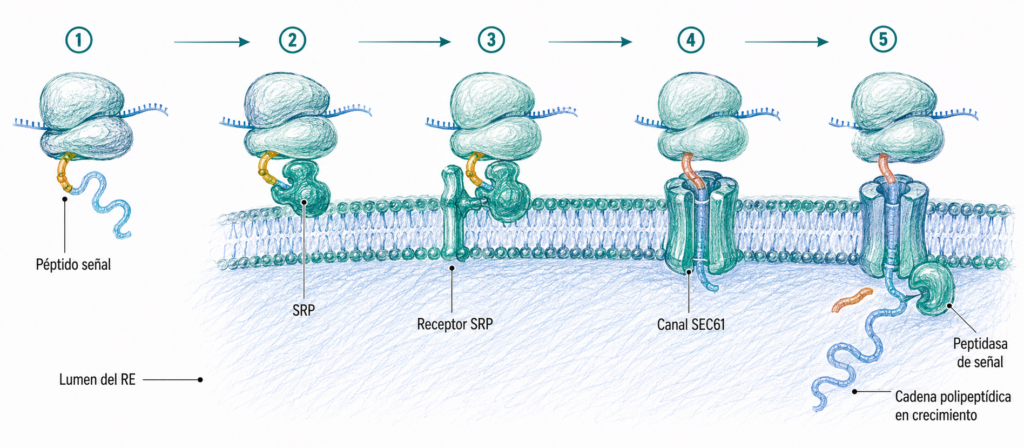
2. Las proteínas transmembrana contienen uno o varios segmentos hidrofóbicos que se anclan en la bicapa lipídica del RE. Estos segmentos detienen la translocación y quedan integrados en la membrana.
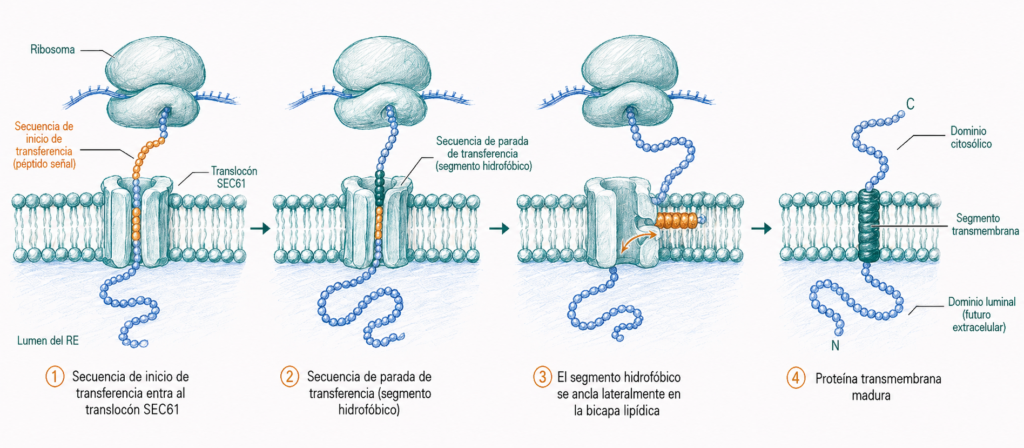
La síntesis cotraduccional garantiza que las proteínas de la vía secretora nunca entren en contacto con el citoplasma: se sintetizan directamente en el lumen del RE o se integran en su membrana.
6.3.2. Glicosilación de proteínas en el RE
La mayoría de las proteínas que entran al lumen del RER son glicosiladas durante o inmediatamente después de la translocación. Este proceso se denomina N-glicosilación porque el oligosacárido se une al nitrógeno del grupo amino de un residuo de asparagina (Asn) en la secuencia consenso Asn-X-Ser/Thr.
El proceso es el siguiente: un oligosacárido precursor formado por 14 residuos de azúcar (2 N-acetilglucosaminas + 9 manosas + 3 glucosas) se ensambla en la membrana del RE unido a una molécula lipídica denominada dolicol. El complejo dolicol-oligosacárido se sitúa inicialmente en la hemimembrana citosólica, donde se añaden los primeros azúcares; a continuación, una flipasa (escramblasa) lo transloca al lumen, donde se completa la síntesis del oligosacárido.
La enzima oligosacariltransferasa transfiere el oligosacárido completo desde el dolicol al residuo de asparagina de la proteína en síntesis.
Inmediatamente después, el oligosacárido comienza a ser modificado en el propio lumen del RE: se eliminan las 3 glucosas y 1 manosa antes de que la proteína abandone el RE. Este procesamiento inicial tiene una función de control de calidad, como se describe a continuación.
6.3.3. Control de calidad: degradación de proteínas mal plegadas (ERAD)
El RE no permite que las proteínas mal plegadas o incorrectamente ensambladas avancen por la vía secretora. Las chaperonas del lumen del RE (como la BiP/GRP78, homóloga de la Hsp70) reconocen y retienen las proteínas con estructuras incorrectas.
El estado de glicosilación actúa como indicador de plegamiento. Las proteínas recién glicosiladas tienen tres glucosas en el oligosacárido; a medida que se pliegan correctamente, las glucosas se van eliminando. Si la proteína está mal plegada, una glucosiltransferasa reañade una glucosa al oligosacárido, haciendo que la proteína vuelva a ser retenida por las chaperonas en un ciclo de control.
Las proteínas que no consiguen plegarse correctamente son exportadas de vuelta al citoplasma mediante el proceso inverso a la translocación (retrotranslocación o dislocación). Antes de la ubiquitinación, la N-glicanasa elimina los oligosacáridos de la proteína retrotranslocada, facilitando su reconocimiento por el sistema ubiquitina-proteasoma. En el citoplasma son ubiquitinadas y degradadas por el proteasoma. Este proceso se denomina ERAD (ER-Associated Degradation).
Cuando la demanda de síntesis de proteínas supera la capacidad de plegamiento del RE, se acumulan proteínas mal plegadas en el lumen. Esto activa la respuesta a proteínas desplegadas (UPR), que intenta restaurar la homeostasis aumentando la capacidad del RE o, si el estrés es irreversible, activando la apoptosis. La UPR está implicada en la patogénesis de enfermedades tan diversas como la diabetes tipo 2 (células β pancreáticas), enfermedades neurodegenerativas y algunos cánceres.
6.4. Funciones del RE liso
6.4.1. Síntesis de lípidos
El REL es el principal sitio de síntesis de lípidos de la célula. En su membrana se localizan las enzimas responsables de la síntesis de fosfolípidos, ceramida y colesterol, que son los lípidos de las membranas de todos los orgánulos y de la membrana plasmática.
La síntesis ocurre en la hemimembrana citosólica del REL: las enzimas tienen sus sitios activos orientados hacia el citoplasma y los lípidos sintetizados se incorporan inicialmente a esta hemimembrana. Las flipasas del RE redistribuyen los lípidos entre las dos hemimembranas para mantener el equilibrio.
Desde el REL, los lípidos se distribuyen al resto de membranas celulares por dos vías: mediante vesículas de transporte hacia el aparato de Golgi y la membrana plasmática, y mediante proteínas transportadoras de lípidos hacia las mitocondrias y los peroxisomas.
6.4.2. Detoxificación
El REL hepático contiene el sistema enzimático de detoxificación más importante del organismo. Las sustancias tóxicas, habitualmente liposolubles, son transformadas en moléculas más hidrosolubles que pueden excretarse por la orina o la bilis.
La familia del citocromo P450 es el componente central de este sistema. Son enzimas de la membrana del REL que catalizan reacciones de oxidación de sustancias tóxicas endógenas (bilirrubina, hormonas) y exógenas (fármacos, contaminantes, carcinógenos). Las reacciones de oxidación aumentan la hidrosolubilidad de las moléculas, facilitando su eliminación.
El citocromo P450 hepático es también responsable del metabolismo de la mayoría de los fármacos. Las variaciones genéticas en los genes CYP (que codifican las distintas isoformas del P450) explican las diferencias interindividuales en la respuesta a los fármacos: metabolizadores rápidos, lentos o ultrarrápidos. Esta variabilidad tiene implicaciones directas en la dosificación y la toxicidad farmacológica.
6.4.3. Almacén de calcio
El lumen del RE contiene grandes cantidades de Ca²⁺, retenido por proteínas fijadoras de calcio como la calnexina y la calreticulina. El RE es el principal reservorio de calcio intracelular.
En respuesta a señales extracelulares que generan el segundo mensajero IP₃ (inositol trifosfato), los canales de Ca²⁺ del RE (receptores de IP₃) se abren y liberan Ca²⁺ al citoplasma. Este aumento brusco de Ca²⁺ citosólico desencadena múltiples respuestas: contracción muscular, secreción, activación enzimática y cambios en la expresión génica.
El Ca²⁺ es posteriormente recapturado hacia el lumen del RE mediante bombas de Ca²⁺ ATPasa (SERCA).
6.5. El RE como vía de transporte intracelular
El RE es el punto de partida de la vía secretora: la ruta que siguen las proteínas y lípidos desde su síntesis hasta su destino final (secreción al exterior, incorporación a membranas o almacenamiento en orgánulos).
Las proteínas correctamente plegadas y glicosiladas salen del RE en vesículas de transporte que brotan de las zonas del elemento transicional. Estas vesículas tienen una cubierta proteica específica (COP II) que selecciona las proteínas a transportar y dirige las vesículas hacia el aparato de Golgi.
Las vesículas solo pueden fusionarse con la membrana diana correcta: el proceso está controlado por proteínas SNARE que garantizan la especificidad del destino. Proteínas residentes del RE que escapan accidentalmente en vesículas son recapturadas desde el Golgi por vesículas retrógranas recubiertas de COP I, gracias a secuencias de retención (KDEL en proteínas solubles, KKXX en proteínas transmembrana).
El transporte vesicular desde el RE, el papel del aparato de Golgi en la modificación y clasificación de proteínas, y los mecanismos de formación de vesículas se desarrollan en T7 — El aparato de Golgi.
6.6. Biogénesis del RE
El RE no se forma de nuevo en cada ciclo celular: crece a partir del RE preexistente y se divide durante la mitosis para que cada célula hija reciba su propia red de RE.
Los lípidos de la membrana del RE se sintetizan en el propio RE liso. Las proteínas del lumen se sintetizan en el RER y se vierten al lumen directamente. Las proteínas de membrana del RE se sintetizan también en el RER y se integran en su membrana.
Durante la mitosis, la envoltura nuclear se fragmenta y se integra en el RE, que queda distribuido entre las dos células hijas. Al final de la mitosis, el RE vuelve a diferenciarse en envoltura nuclear y RE citoplasmático en cada célula hija.
