7.1. Descubrimiento y ultraestructura
El aparato de Golgi fue descubierto en 1898 por Camillo Golgi, que impregnó células de cerebelo de lechuza con tetraóxido de osmio y observó una estructura reticular perinuclear a la que denominó «sistema reticular interno». Durante décadas se discutió si era un orgánulo real o un artefacto de tinción; con la llegada del microscopio electrónico de transmisión en los años 50 se confirmó su existencia como orgánulo formado por cisternas y vesículas laminares.
Morfológicamente, el aparato de Golgi está formado por dictiosomas: pilas de cisternas aplanadas con los extremos dilatados, rodeadas de vesículas. Cada dictiosoma tiene entre 4 y 8 cisternas superpuestas. Una célula puede tener desde un único dictiosoma hasta cientos de ellos, pero todos forman parte de un único aparato de Golgi continuo, confirmado con el microscopio electrónico de alto voltaje.
El aparato de Golgi se localiza generalmente cerca del núcleo, próximo al centrosoma. En células muy polarizadas como los hepatocitos puede aparecer como dictiosomas dispersos por el citoplasma.
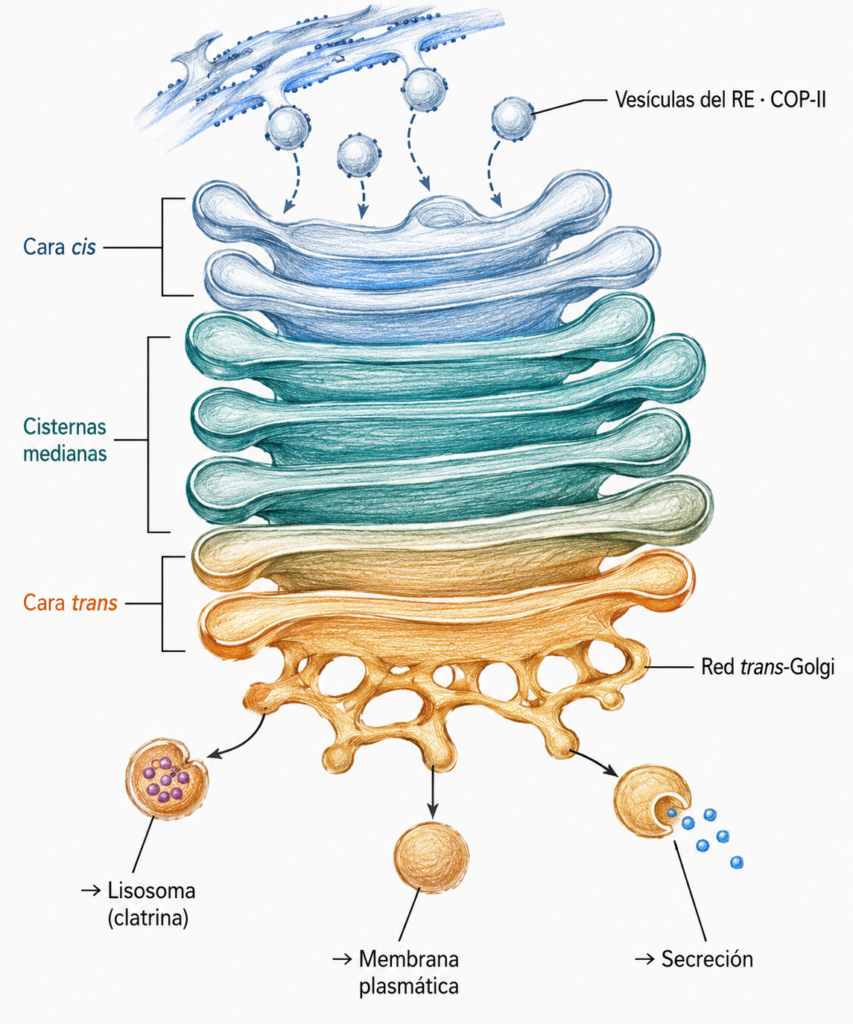
7.2. Polaridad del aparato de Golgi: cara cis y cara trans
El aparato de Golgi está polarizado: sus dos caras tienen composición enzimática, morfología y función distintas.
La cara cis (o de formación) es la que recibe las vesículas procedentes del RE. Está asociada a la red cis-Golgi, una estructura túbulo-vesicular irregular que actúa como punto de entrada. Las vesículas del RE se fusionan entre sí formando el agregado túbulo-vesicular, que después contacta con la red cis-Golgi.
La cara trans (o de secreción/maduración) es la que origina las vesículas de salida hacia los destinos finales. Está asociada a la red trans-Golgi, la cisterna más heterogénea y compleja del aparato, desde donde parten las vesículas hacia la membrana plasmática, los lisosomas o los gránulos de secreción.
Entre la cara cis y la cara trans se sitúan las cisternas medianas, en número variable según el tipo celular, donde ocurren la mayoría de las modificaciones de los azúcares.
El recorrido de una proteína a través del Golgi va siempre en dirección cis→trans. Cada cisterna tiene un conjunto específico de enzimas que realizan modificaciones secuenciales: si no ocurre la primera reacción, no pueden ocurrir las siguientes.
7.3. Funciones del aparato de Golgi
7.3.1. Modificación de proteínas: glicosilación y sulfatación
La función modificadora principal del Golgi es la maduración del oligosacárido iniciada en el RE. Las proteínas llegan al Golgi con un oligosacárido de alto contenido en manosa (2 N-acetilglucosaminas y múltiples manosas). En cada cisterna, enzimas específicas modifican progresivamente este oligosacárido:
- En la cisterna cis, la manosidasa tipo I elimina manosas.
- En las cisternas medianas, se añaden residuos de N-acetilglucosamina y se eliminan más manosas.
- En las cisternas trans, se añaden galactosa y finalmente ácido siálico (NANA), que confiere carga negativa a la proteína.
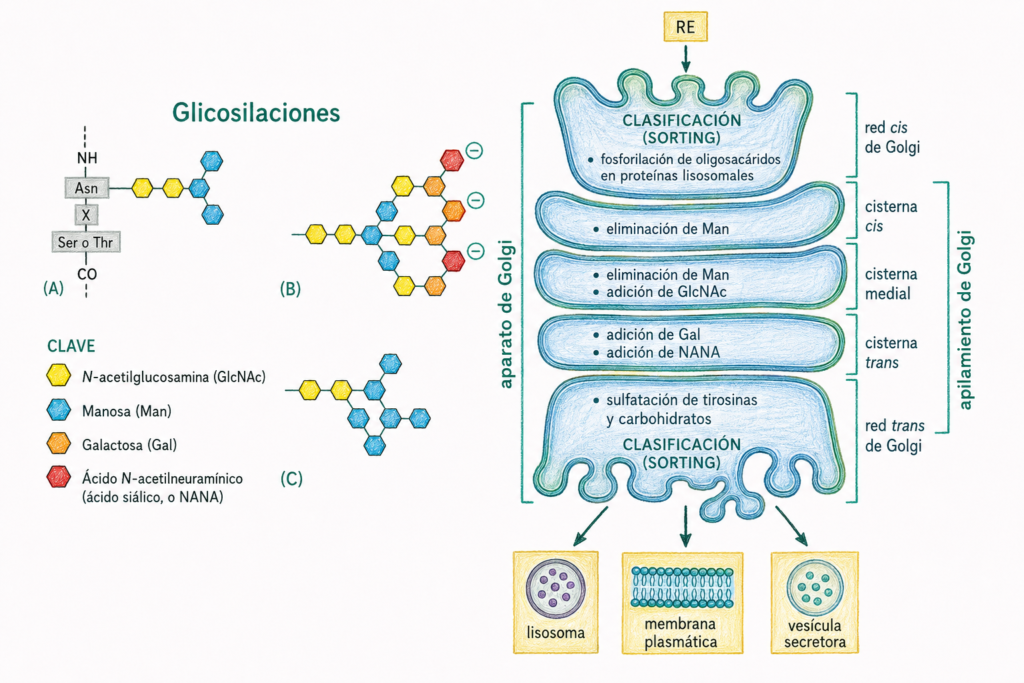
El resultado final depende de la accesibilidad del oligosacárido: los que quedan expuestos en la superficie de la proteína son modificados; los que quedan en el interior de la proteína mantienen su estructura rica en manosa.
| Cisterna | Enzimas presentes | Modificación realizada | Resultado |
|---|---|---|---|
| Red cis-Golgi | Manosidasa tipo I | Elimina manosas del oligosacárido de alto contenido en manosa | Oligosacárido con menos manosas |
| Cisternas medianas (cis→trans) | N-acetilglucosaminiltransferasa I y II · manosidasa tipo II | Añade GlcNAc · elimina más manosas | Oligosacárido híbrido |
| Cisternas medianas tardías | Fucosiltransferasa | Añade fucosa al núcleo del oligosacárido | Oligosacárido complejo con fucosa |
| Cisternas trans | Galactosiltransferasa | Añade galactosa | Cadenas con galactosa terminal |
| Red trans-Golgi | Sialiltransferasa · sulfotransferasa | Añade ácido siálico (carga negativa) · sulfata residuos de Tyr | Oligosacárido complejo maduro con carga negativa |
Todas las glucoproteínas comparten una región común: 2 N-acetilglucosaminas y 3 manosas. A partir de esta base común se construyen los oligosacáridos complejos específicos de cada proteína.
El Golgi también realiza sulfatación de proteínas en la red trans-Golgi: sulfotransferasas añaden grupos sulfato, principalmente a residuos de tirosina de proteínas destinadas a la secreción. El proceso requiere la activación previa del sulfato por ATP.
Además de las N-glicosilaciones iniciadas en el RE, en el Golgi se producen las O-glicosilaciones: adición de azúcares a residuos de serina y treonina de proteínas. Las O-glicosilaciones son frecuentes en proteoglicanos y mucinas.
7.3.2. Clasificación y envío a destinos
Desde la red trans-Golgi, las proteínas se clasifican y se envían a tres destinos principales:
- Lisosomas: las enzimas lisosomales son marcadas en el Golgi con manosa-6-fosfato (M6P). Este marcador es reconocido por receptores específicos de M6P en la membrana del Golgi, que concentran las enzimas lisosomales en vesículas recubiertas de clatrina destinadas a los lisosomas.
- Membrana plasmática y secreción al exterior: por las vías de secreción constitutiva y regulada (ver 7.3.3).
- Reciclaje al RE: las proteínas residentes del RE que escapan accidentalmente son recuperadas desde el Golgi por vesículas COP-I y devueltas al RE (ver 7.4.2).
7.3.3. Secreción constitutiva y regulada
La secreción constitutiva es la vía por defecto: todas las proteínas que no tienen ninguna señal específica de destino salen del Golgi en vesículas que se fusionan continuamente con la membrana plasmática. Es una vía presente en todos los tipos celulares. Por esta vía salen componentes del glicocáliz, proteínas de la matriz extracelular y lípidos de membrana (incluyendo los lipid rafts).
La secreción regulada es exclusiva de células especializadas: neuronas, células endocrinas y exocrinas, mastocitos. Las proteínas se acumulan en gránulos de secreción inmaduros que brotan de la red trans-Golgi recubiertos de clatrina. Los gránulos maduran progresivamente: la clatrina se despolimeriza, el interior se acidifica por bombas de protones y las proteínas se condensan hasta 200 veces. Los gránulos maduros permanecen en el citoplasma hasta recibir la señal de secreción (habitualmente un aumento de Ca²⁺ intracelular), momento en que se fusionan con la membrana plasmática y liberan su contenido al exterior.
7.3.4. Reciclaje de membranas
Las vesículas de secreción se fusionan continuamente con la membrana plasmática, aportándole nuevos lípidos y proteínas. Este aumento de superficie membranaria se compensa con la formación simultánea de vesículas de endocitosis, que recuperan membrana del interior. El balance entre exocitosis y endocitosis mantiene constante el área total de la membrana plasmática.
7.4. Transporte vesicular
El aparato de Golgi es una estructura dinámica en constante renovación de vesículas. Las cisternas están rodeadas de vesículas en todo momento. El transporte entre compartimentos es altamente específico: cada tipo de vesícula solo puede fusionarse con su membrana diana, gracias al sistema de proteínas SNARE que garantiza la especificidad del destino.
Las vesículas de transporte se clasifican según su cubierta proteica:
| Tipo | Cubierta | Dirección | Origen → Destino | Función principal |
|---|---|---|---|---|
| COP-II | Proteínas Sec23/24/13/31 | Anterógrado | RE → Golgi (red cis) | Transporte de proteínas recién sintetizadas del RE al Golgi |
| COP-I | Proteínas COP (α β γ δ ε ζ) | Retrógrado | Golgi → RE · entre cisternas Golgi | Recuperación de proteínas residentes del RE · reciclaje entre cisternas |
| Clatrina | Triskeliones de clatrina + adaptinas | Variable | Red trans-Golgi → lisosomas · endosomas → membrana | Transporte de enzimas lisosomales (M6P) · endocitosis mediada por receptores |
7.4.1. Vesículas recubiertas de clatrina
La clatrina es una proteína estructural que forma una red polimérica en la cara citosólica de la membrana. Sus unidades básicas son los triskeliones: monómeros formados por 3 cadenas pesadas y 3 cadenas ligeras. Tres triskeliones se unen formando estructuras de hexágonos y pentágonos que recubren la vesícula en formación.
La formación de la vesícula de clatrina requiere proteínas accesorias: las adaptinas (que seleccionan las proteínas cargo y ligan la clatrina a la membrana) y la dinamina (que contrae el cuello de la vesícula y la separa de la membrana).
Las vesículas de clatrina median el transporte entre el Golgi y los lisosomas (llevando enzimas lisosomales marcadas con M6P) y entre el Golgi y la membrana plasmática en la secreción regulada.
7.4.2. Vesículas COP-I y COP-II
Las vesículas COP-II median el transporte anterógrado desde el RE hacia el Golgi. Se forman en el elemento transicional del RE, son muy selectivas en las proteínas que capturan y se fusionan con la red cis-Golgi.
Las vesículas COP-I median el transporte retrógrado: desde el Golgi de vuelta al RE y entre las cisternas del Golgi. Son responsables de recuperar las proteínas residentes del RE que han escapado accidentalmente: reconocen las secuencias de retención (KDEL en proteínas solubles, KKXX en proteínas transmembrana) y las devuelven al RE.
El transporte vesicular anterógrado (RE→Golgi→membrana) está asociado a los microtúbulos: cuando se despolimerizan con colchicina, el aparato de Golgi se fragmenta y dispersa.
7.5. Biogénesis del aparato de Golgi
La biogénesis del Golgi no está completamente resuelta. Existen dos modelos no excluyentes:
El modelo de maduración de cisternas propone que las cisternas no son estructuras estables sino que se forman en la cara cis a partir de vesículas del RE, maduran progresivamente adquiriendo las enzimas características de cada cisterna y finalmente se consumen en la cara trans. Las enzimas de cada cisterna se mantienen en su posición gracias al transporte retrógrado por vesículas COP-I.
El modelo de transporte vesicular propone que las cisternas son estructuras estables y las proteínas viajan entre ellas en vesículas.
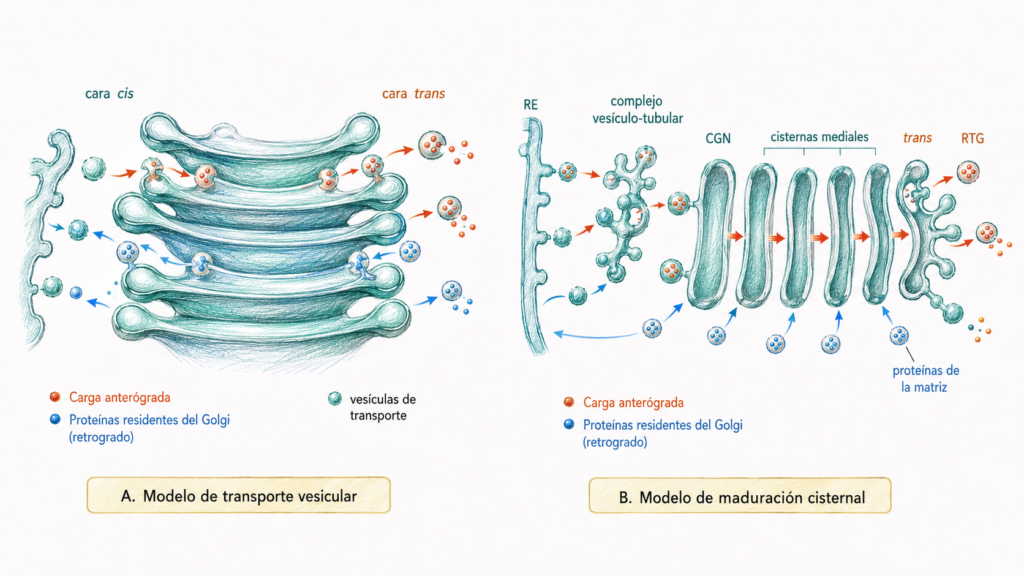
La evidencia actual apoya principalmente el modelo de maduración, aunque el transporte vesicular también contribuye.
Las proteínas de la matriz del Golgi (como GM130 y p115) estabilizan la estructura de las cisternas y las mantienen apiladas. Al inicio de la mitosis, estas proteínas se fosforilan, el Golgi se fragmenta en vesículas que se distribuyen entre las células hijas, y al terminar la división se desfosforilan y el Golgi se reconstituye.
La biogénesis de los lisosomas a partir del aparato de Golgi, el papel del marcador manosa-6-fosfato y las enfermedades lisosomales se desarrollan en T8 — El lisosoma.
