10.1. Introducción al citoesqueleto
El citoesqueleto es una red tridimensional de filamentos proteicos que ocupa el citoplasma de las células eucariotas. Es exclusivo de las eucariotas: las células procariotas no tienen citoesqueleto equivalente.
Sus funciones principales son el mantenimiento de la forma celular, el movimiento de la propia célula, el transporte y posicionamiento de orgánulos y vesículas, y la organización del huso mitótico durante la división celular.
El citoesqueleto está formado por tres tipos de filamentos proteicos, cada uno con propiedades mecánicas y funciones distintas:
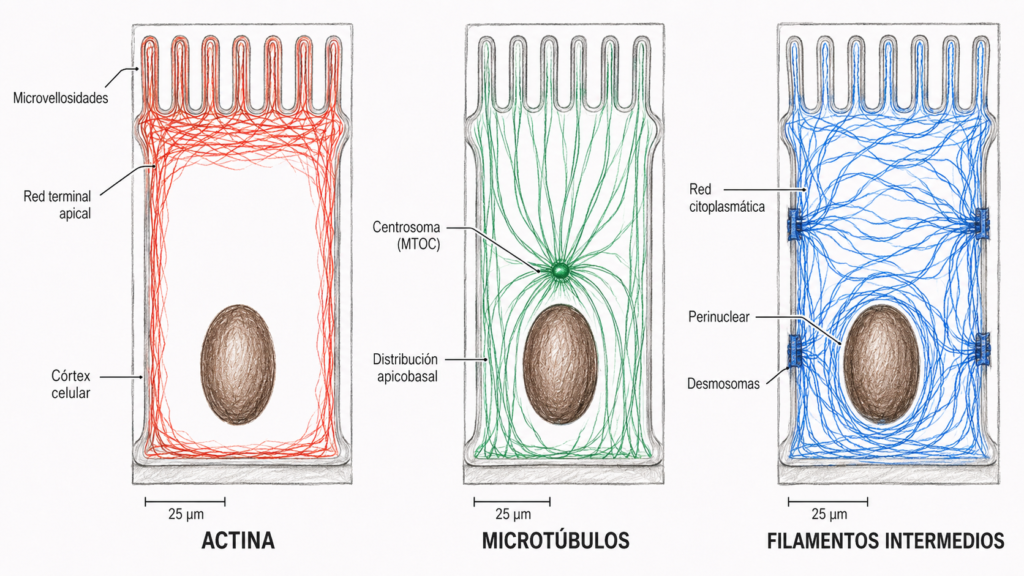
- Microtúbulos: cilindros huecos de 25 nm de diámetro formados por tubulina. Son los más rígidos y los responsables del movimiento de estructuras intracelulares.
- Microfilamentos de actina: filamentos de 7 nm de diámetro formados por actina. Son los más delgados y responsables de la forma de la superficie celular, la motilidad y la contracción.
- Filamentos intermedios: filamentos de 10-11 nm de diámetro formados por diversas proteínas fibrosas. Son los más resistentes mecánicamente y proporcionan soporte estructural.
| Característica | Microtúbulos | Microfilamentos de actina | Filamentos intermedios |
|---|---|---|---|
| Diámetro | 25 nm | 7 nm | 10-11 nm |
| Proteína principal | Tubulina α/β (heterodímero) | Actina (G-actina → F-actina) | Variable según tipo celular (queratina, vimentina, desmina...) |
| Polaridad | Sí (extremo + y -) | Sí (extremo barba + y punta -) | No |
| Dinámica | Alta (inestabilidad dinámica · catástrofes) | Alta (treadmilling) | Baja (más estables) |
| Proteínas motoras | Kinesina (→+) · dineína (→-) | Miosina (→+) | No |
| Función principal | Transporte intracelular · huso mitótico · cilios | Forma celular · motilidad · citocinesis · contracción | Soporte mecánico · resistencia al estrés |
| Inhibidores farmacológicos | Colchicina (despolimeriza) · taxol (estabiliza) | Citocalasina (despolimeriza) · falolidina (estabiliza) | No hay fármacos específicos |
Los tres tipos comparten características comunes: son filamentos proteicos formados por monómeros que se polimerizan mediante fuerzas no covalentes débiles, y sus funciones dependen tanto de su estructura como de las proteínas asociadas que se unen a ellos.
10.2. Microtúbulos: morfología y composición
Los microtúbulos son cilindros huecos de 25 nm de diámetro exterior y 5 nm de grosor de pared. Su longitud es muy variable: pueden aparecer y desaparecer rápidamente en distintas localizaciones de la célula.
Están formados por 13 protofilamentos dispuestos en paralelo alrededor del eje del cilindro. Cada protofilamento es una cadena de heterodímeros de tubulina: la subunidad α-tubulina y la β-tubulina, ambas de ~55 kDa con estructura globular muy similar. En mamíferos existen seis isoformas de cada tipo, codificadas por genes distintos.
Los heterodímeros tienen sitios de unión para GTP, GDP, colchicina y alcaloides (vincristina, vinblastina), que son las drogas que interfieren con la dinámica de los microtúbulos.
Los microtúbulos son estructuras polares: sus dos extremos tienen propiedades distintas:
- El extremo (+) crece rápidamente y apunta hacia la periferia celular.
- El extremo (-) crece lentamente y está generalmente anclado en el centrosoma.
10.3. Dinámica de los microtúbulos: polimerización y catástrofe
Los microtúbulos son estructuras dinámicas en equilibrio constante entre polimerización y despolimerización. En una célula en interfase, aproximadamente la mitad de la tubulina está polimerizada y la otra mitad está libre en el citoplasma.
La polimerización se inicia por nucleación en el centrosoma. Los heterodímeros α/β con GTP unido se incorporan al extremo (+) del microtúbulo en crecimiento. Mientras el microtúbulo crece, el GTP de la β-tubulina se hidroliza a GDP en el interior del filamento. El extremo (+) en crecimiento mantiene un casquete de GTP que estabiliza el microtúbulo.
Cuando la hidrólisis de GTP supera la velocidad de adición de nuevos dímeros, el casquete de GTP se pierde y el extremo (+) queda expuesto con GDP-tubulina, que tiene baja afinidad por el microtúbulo. Se produce entonces una despolimerización brusca denominada catástrofe, en la que el microtúbulo se acorta rápidamente hasta desaparecer o hasta que un nuevo casquete de GTP estabiliza el extremo.
Esta propiedad de alternar fases de crecimiento y colapso se denomina inestabilidad dinámica y es esencial para que los microtúbulos exploren el espacio citoplasmático y capturen los cinetocoros de los cromosomas durante la mitosis.
Varios fármacos antitumorales actúan sobre la dinámica de los microtúbulos, impidiendo la formación del huso mitótico y bloqueando la división celular:
- Colchicina y alcaloides de la vinca (vincristina, vinblastina): impiden la polimerización de la tubulina bloqueando el microtúbulo en fase de despolimerización. Usados en gota (colchicina) y en oncología (alcaloides de la vinca).
- Taxol (paclitaxel): estabiliza los microtúbulos impidiendo su despolimerización. Las células no pueden desensamblar el huso mitótico al final de la mitosis. Usado en cáncer de mama, ovario y pulmón.
10.4. El centrosoma: MTOC principal
El centrosoma es el centro organizador de microtúbulos (MTOC) principal de las células animales. Contiene dos centriolos dispuestos perpendicularmente entre sí (formando el diplosoma) rodeados de material pericentriolar (PCM), una matriz proteica amorfa que contiene los componentes necesarios para nucleotar los microtúbulos.
La nucleación de microtúbulos en el centrosoma depende de la tubulina γ, que forma anillos (γ-TuRC, γ-tubulin ring complex) en el PCM. Estos anillos actúan como plantilla para iniciar la polimerización de los 13 protofilamentos del microtúbulo, siempre por el extremo (-). Por eso los microtúbulos irradian desde el centrosoma hacia la periferia con el extremo (+) apuntando hacia afuera.
10.5. Tipos de microtúbulos: lábiles y estables
10.5.1. Microtúbulos lábiles
Los microtúbulos lábiles son los microtúbulos citoplasmáticos dinámicos. Son sensibles a la temperatura (se despolimerizan a 4°C) y a la colchicina. Requieren fijación cuidadosa para visualizarse al microscopio electrónico.
Incluyen los microtúbulos citoplasmáticos que irradian desde el centrosoma en células en interfase, y los microtúbulos del huso mitótico que se ensamblan durante la división celular para segregar los cromosomas.
10.5.2. Microtúbulos estables
Los microtúbulos estables resisten la temperatura baja y los fijadores habituales. Forman parte de estructuras permanentes: cilios, flagelos y centriolos. Su estabilidad se debe a modificaciones post-traduccionales de la tubulina (acetilación, poliglutamilación) y a proteínas asociadas que los estabilizan.
| Característica | Lábiles | Estables |
|---|---|---|
| Sensibilidad al frío | Se despolimerizan a 4°C | Resisten temperaturas bajas |
| Sensibilidad a colchicina | Sí | No (o muy poco) |
| Modificaciones de tubulina | Escasas | Acetilación · poliglutamilación · detirosinalización |
| Dinámica | Alta (inestabilidad dinámica) | Baja |
| Localización | Microtúbulos citoplasmáticos · huso mitótico | Cilios · flagelos · centriolos |
| Función | Transporte vesicular · segregación cromosómica | Movimiento ciliar · nucleación de microtúbulos |
10.6. Cilios y flagelos
Los cilios y los flagelos son extensiones citoplasmáticas móviles que realizan movimientos pendulares u ondulantes. Tienen el mismo diámetro (~0,25 μm) y la misma estructura interna, pero difieren en longitud y número: los cilios miden ~10 μm y son numerosos; los flagelos miden ~50 μm y generalmente son uno o pocos por célula.
Funcionalmente también difieren: los cilios mueven el líquido extracelular sobre la superficie celular (epitelio respiratorio, trompas de Falopio) mientras que los flagelos mueven la célula en su conjunto (espermatozoide).
10.6.1. Estructura del axonema: 9+2
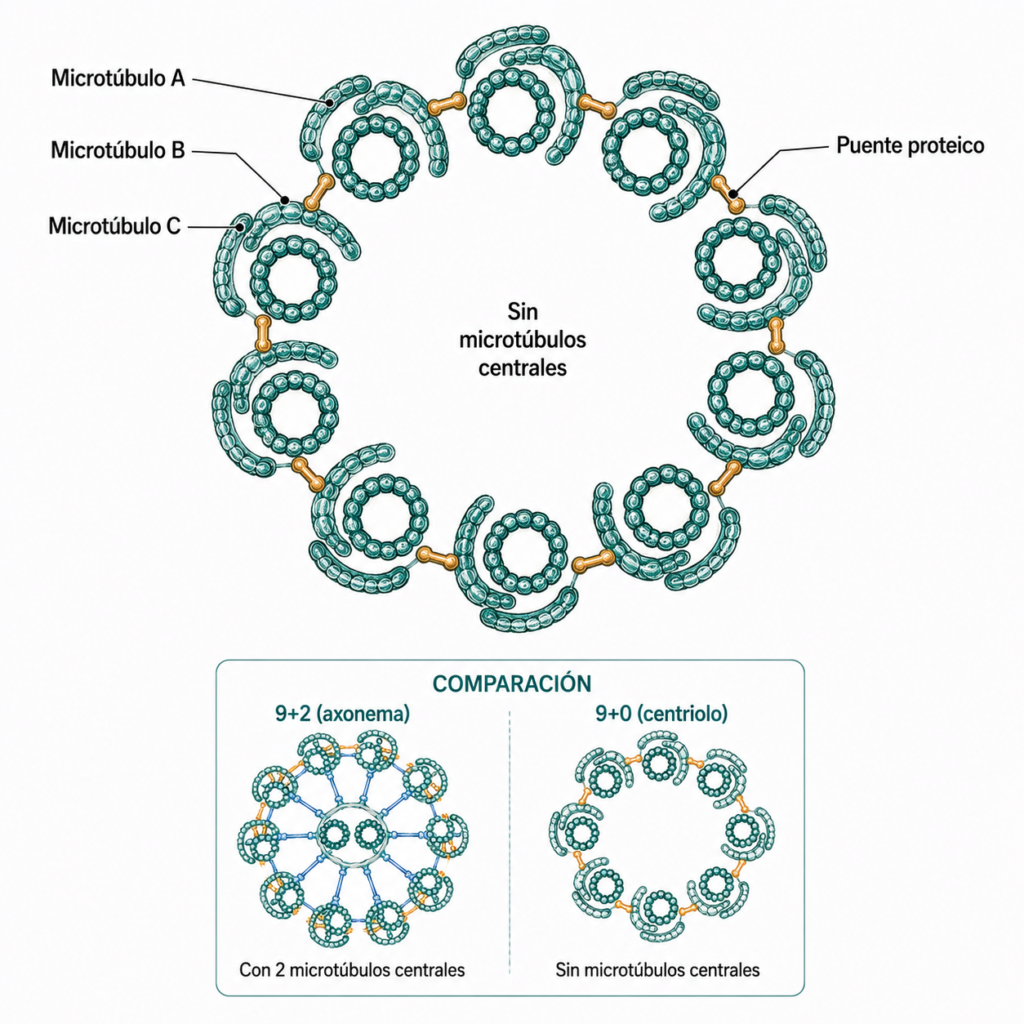
El esqueleto interno del cilio o flagelo se denomina axonema y tiene una estructura conservada en prácticamente todos los eucariotas: 9 dobletes de microtúbulos periféricos + 2 microtúbulos centrales (estructura 9+2).
Cada doblete periférico está formado por dos microtúbulos fusionados: el microtúbulo A (sección circular completa, 13 protofilamentos) y el microtúbulo B (sección de media luna, 10-11 protofilamentos, fusionado al A).
Los dobletes adyacentes están unidos por puentes de nexina, proteínas elásticas que mantienen la integridad del axonema durante el movimiento.
Del microtúbulo A de cada doblete surgen dos tipos de proyecciones proteicas: los brazos de dineína externos e internos hacia el doblete vecino, y las fibras radiales que apuntan hacia la vaina central que rodea los dos microtúbulos centrales.
10.6.2. Mecanismo del movimiento ciliar
El movimiento ciliar se genera por el deslizamiento de los dobletes entre sí, no por su acortamiento. Los brazos de dineína son ATPasas (dineína axonemal) que utilizan la energía de la hidrólisis de ATP para desplazarse a lo largo del microtúbulo B del doblete vecino en dirección al extremo (-).
Como los dobletes están unidos entre sí por los puentes de nexina y anclados en el corpúsculo basal, el deslizamiento se convierte en una flexión del axonema completo. La regulación coordinada de los brazos de dineína en distintas posiciones del axonema genera el patrón de movimiento característico: batido pendular en los cilios, movimiento ondulante en los flagelos.
10.7. Centriolos y corpúsculos basales
Los centriolos y los corpúsculos basales (cinetosomas) tienen la misma estructura y son funcionalmente intercambiables según su posición en la célula.
Su estructura es 9+0: 9 tripletes de microtúbulos dispuestos en anillo, sin microtúbulos centrales. Cada triplete tiene tres microtúbulos: A (sección circular), B (media luna, fusionado al A) y C (media luna, fusionado al B). Los tripletes están unidos entre sí por puentes proteicos entre el microtúbulo A de un triplete y el C del siguiente.
Miden ~0,2 μm de diámetro y ~0,4 μm de longitud. En la zona proximal presentan una estructura proteica en forma de rueda de carro (hub central con 9 radios) no presente en la zona distal.
Las células animales tienen al menos dos centriolos perpendiculares entre sí formando el diplosoma en el centrosoma. Las células ciliadas tienen un corpúsculo basal anclando cada cilio, conectado a la base del axonema por una placa basal densa.
Los centriolos del centrosoma pueden convertirse en corpúsculos basales para nucleotar cilios (por ejemplo, durante la diferenciación del epitelio respiratorio). Inversamente, los corpúsculos basales pueden actuar como centrosoma al inicio de la mitosis en algunas células ciliadas.
La discinesia ciliar primaria es un grupo de enfermedades genéticas causadas por defectos en las proteínas del axonema, más frecuentemente en los brazos de dineína. Los cilios están presentes pero inmóviles o con movimiento anómalo.
Las consecuencias clínicas derivan de la pérdida de función ciliar en distintos órganos: infecciones respiratorias de repetición (bronquiectasias, sinusitis crónica por aclaramiento mucociliar deficiente), infertilidad masculina (espermatozoides inmóviles) e hidrocefalia (circulación deficiente del líquido cefalorraquídeo).
El síndrome de Kartagener es la variante más conocida: además de los síntomas anteriores, presenta situs inversus (transposición de órganos). Durante el desarrollo embrionario, los cilios del nodo embrionario establecen la asimetría izquierda-derecha mediante su movimiento rotacional; sin cilios funcionales, la lateralización ocurre al azar, resultando en situs inversus en aproximadamente el 50% de los casos.
