11.1. Microfilamentos de actina
11.1.1. Estructura: G-actina y F-actina
La actina es la proteína más abundante de las células eucariotas no musculares, representando hasta el 10-15% de la proteína total. Existen seis isoformas en mamíferos: α (músculo), β y γ (no muscular), con pequeñas diferencias de secuencia.
La unidad básica es la G-actina (actina globular): un monómero de ~42 kDa con forma esférica que tiene un sitio de unión a ATP y a cationes divalentes (Ca²⁺ o Mg²⁺). Los monómeros de G-actina se polimerizan formando el filamento de actina o F-actina: una hélice de doble cadena con un diámetro de ~7 nm.
Los filamentos de actina son polares: sus dos extremos tienen propiedades cinéticas distintas. El extremo barba o extremo (+) tiene mayor velocidad de polimerización. El extremo punta o extremo (-) polimeriza más lentamente y despolimeriza con mayor facilidad.
11.1.2. Dinámica de polimerización
La polimerización de la actina ocurre en tres fases:
- Nucleación: formación de un oligómero estable de 3-4 monómeros que actúa como semilla. Es el paso limitante — los dímeros y trímeros son inestables.
- Elongación: adición rápida de monómeros G-actina-ATP en el extremo (+). A medida que la actina se incorpora al filamento, el ATP se hidroliza a ADP, produciendo F-actina-ADP en el interior del filamento.
- Estado estacionario (treadmilling): cuando la concentración de G-actina libre se iguala con la concentración crítica, el filamento alcanza un estado dinámico en el que la polimerización en el extremo (+) y la despolimerización en el extremo (-) ocurren a la misma velocidad. El filamento mantiene su longitud pero sus subunidades se desplazan continuamente desde el extremo (+) al (-): es el fenómeno de treadmilling o pisada de molino.
11.1.3. Proteínas reguladoras de la actina
La dinámica de la actina está finamente regulada por numerosas proteínas accesorias:
- Timosina β4: secuestra monómeros de G-actina en el citoplasma impidiendo su polimerización. Mantiene el reservorio de actina libre disponible para polimerizaciones rápidas.
- Profilina: se une a monómeros de G-actina y facilita su incorporación al extremo (+) de los filamentos. Compite con la timosina: cuando la profilina se activa por señales intracelulares, libera la actina secuestrada por la timosina y favorece la polimerización localizada.
- Complejo ARP2/3: nucleotida la formación de ramas de actina en ángulo de 70° a partir de filamentos preexistentes. Genera redes ramificadas características de los lamelipodios y de otras estructuras de motilidad.
- Cofilina (ADF): se une a F-actina-ADP y favorece la despolimerización del extremo (-), aumentando el recambio dinámico de los filamentos. Esencial para mantener el treadmilling.
- Tropomiosina: se une lateralmente a los filamentos de actina estabilizándolos e impidiendo que interaccionen con otras proteínas. Clave en la regulación de la contracción muscular.
- Fimbrina, α-actinina, filamina: proteínas de entrecruzamiento que organizan los filamentos en haces paralelos (fimbrina, α-actinina) o redes tridimensionales (filamina).
11.1.4. Estructuras de actina en la célula
Los microfilamentos de actina se organizan en distintas estructuras según las proteínas accesorias presentes:
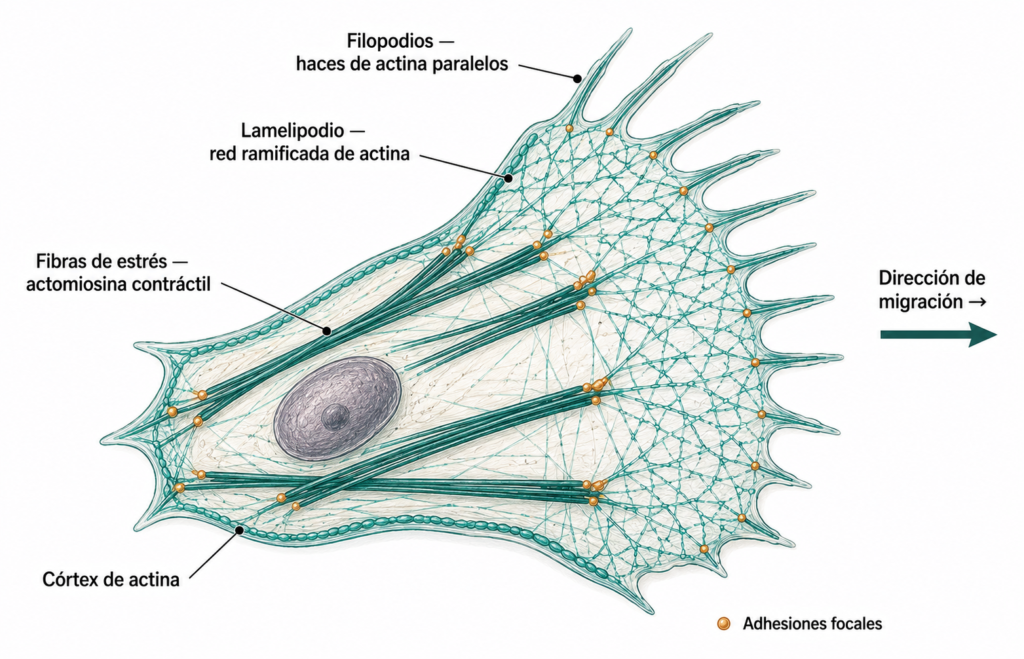
- Córtex celular: red de actina justo debajo de la membrana plasmática que mantiene la forma celular y la rigidez superficial. Está unido a la membrana mediante proteínas de la familia ERM (ezrina, radixina, moesina) y por la espectrina en los eritrocitos.
- Filopodios: haces paralelos de actina muy juntos (estabilizados por fimbrina) que forman proyecciones digitiformes rígidas que exploran el entorno. No son contráctiles.
- Lamelipodios: redes ramificadas de actina (generadas por el complejo ARP2/3) que forman velos planos en el frente de avance de las células en migración.
- Fibras de estrés: haces de actina y miosina II con capacidad contráctil que generan tensión en la célula y se anclan a la matriz extracelular mediante las uniones focales (integrinas).
- Anillo contráctil: estructura de actina y miosina II que se forma en el ecuador celular durante la citocinesis y se contrae para dividir la célula en dos.
11.1.5. Estereocilios: mecanosensores auditivos
Los estereocilios son extensiones de la membrana plasmática de las células ciliadas del oído interno (órgano de Corti). A pesar de su nombre, no son cilios verdaderos: no contienen microtúbulos ni tienen movimiento activo propio. Son proyecciones rígidas con un haz de actina en su interior, estabilizadas por fimbrina y esprina, estructuralmente similares a las microvellosidades pero mucho más largas y ordenadas por tamaño.
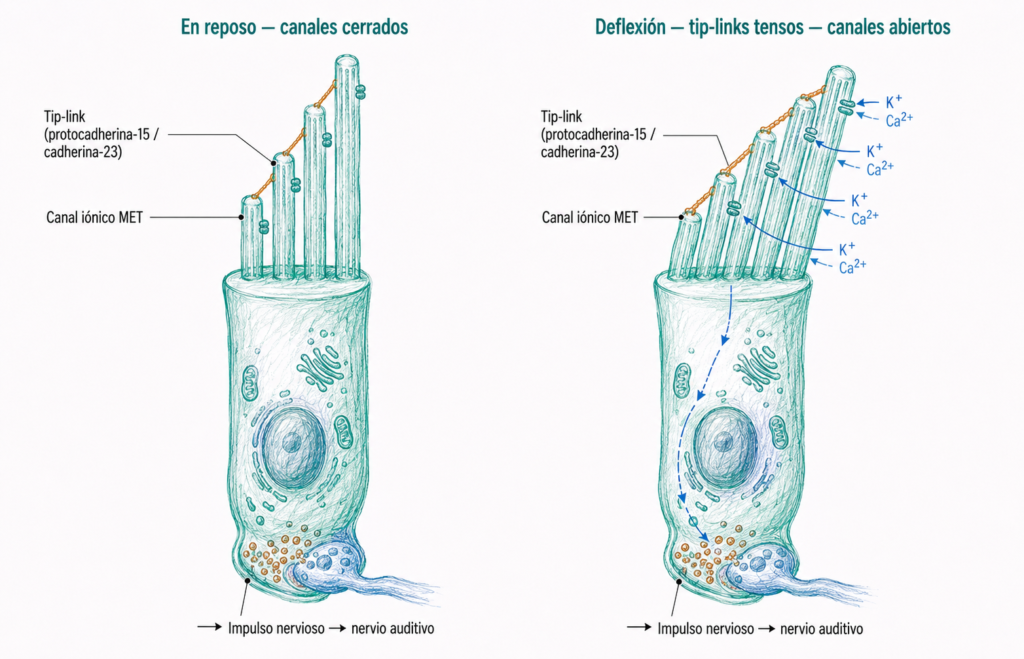
Los estereocilios de una misma célula ciliada están organizados en filas de altura creciente. Entre el extremo de cada estereocilio y la cara lateral del adyacente más alto existe un filamento proteico denominado tip-link (enlace de punta), formado por cadherinas atípicas (protocadherina-15 y cadherina-23).
Cuando el sonido llega al oído interno, las ondas de presión hacen vibrar la membrana basilar, lo que inclina el haz de estereocilios. Esta inclinación tensa los tip-links, que tiran mecánicamente de los canales iónicos de la membrana (canales MET, de mecanotransducción). La apertura de estos canales permite la entrada de K⁺ y Ca²⁺ al interior de la célula ciliada, despolarizando la membrana y generando el impulso nervioso que se transmite por el nervio auditivo.
Las mutaciones en la protocadherina-15 y la cadherina-23 (proteínas de los tip-links) producen el síndrome de Usher, la causa más frecuente de sordoceguera congénita hereditaria. Las mutaciones en la miosina VII, que ancla los estereocilios a la membrana y mantiene los tip-links en tensión, producen también sordera congénita. La miosina VII es el único miembro de la superfamilia de miosinas que se desplaza hacia el extremo (-) de los microfilamentos de actina.
11.2. Filamentos intermedios
11.2.1. Características generales
Los filamentos intermedios tienen un diámetro de 10-11 nm, intermedio entre microfilamentos (7 nm) y microtúbulos (25 nm). Son los componentes más resistentes mecánicamente del citoesqueleto: proporcionan soporte estructural a la célula y resistencia al estrés mecánico.
A diferencia de microtúbulos y microfilamentos, los filamentos intermedios tienen características únicas:
- No son polares: no tienen extremo (+) ni (-). Sus subunidades pueden añadirse en cualquier punto del filamento.
- No son motores: no hay proteínas motoras que se desplacen sobre ellos (a diferencia de miosina sobre actina o dineína/kinesina sobre microtúbulos).
- Son más estables: no tienen la dinámica de polimerización/despolimerización rápida de microtúbulos y microfilamentos, aunque pueden reorganizarse durante la mitosis por fosforilación.
- Son heterogéneos: a diferencia de la tubulina y la actina, los filamentos intermedios están formados por proteínas muy diversas, específicas de cada tipo celular.
11.2.2. Estructura molecular
Todos los filamentos intermedios comparten una arquitectura similar: un dominio central en α-hélice de ~310 aminoácidos (dominio de varilla) flanqueado por dominios N-terminal y C-terminal globulares variables.
El ensamblaje ocurre en varios pasos: dos monómeros forman un dímero en espiral paralelo; dos dímeros se asocian de forma antiparalela formando un tetrámero; los tetrámeros se apilan lateralmente y longitudinalmente formando el filamento de 10-11 nm. La naturaleza antiparalela del tetrámero explica la ausencia de polaridad.
11.2.3. Clasificación según el tipo celular
Los filamentos intermedios se clasifican en cinco grupos según las proteínas que los forman:
- Queratinas (tipo I y II): son los filamentos intermedios de las células epiteliales. Existen más de 50 queratinas distintas en humanos, expresadas en combinaciones específicas según el tipo de epitelio. Su función es resistir el estrés mecánico. Se anclan a los desmosomas y hemidesmosomas.
- Vimentina (tipo III): presente en células de origen mesenquimal: fibroblastos, células endoteliales, linfocitos, macrófagos. Es el marcador inmunohistoquímico estándar de células mesenquimales.
- Desmina (tipo III): filamento intermedio específico del músculo (esquelético, cardíaco y liso). Conecta los sarcómeros adyacentes y ancla los miofilamentos a la membrana celular.
- Proteína ácida fibrilar glial o GFAP (tipo III): filamento intermedio de los astrocitos del sistema nervioso central. Marcador de diferenciación astroglial.
- Neurofilamentos NF-L, NF-M, NF-H (tipo IV): presentes en las neuronas, especialmente en los axones. Determinan el calibre axonal y, por tanto, la velocidad de conducción nerviosa.
- Laminas A, B y C (tipo V): son los filamentos intermedios del núcleo. Forman la lámina nuclear densa que tapiza la cara interna de la membrana nuclear interna. A diferencia del resto de filamentos intermedios, las laminas son nucleares y se fosforilan al inicio de la mitosis provocando la despolimerización de la lámina densa y la desaparición de la envoltura nuclear.
| Tipo | Proteína(s) | Célula que lo expresa | Función principal | Patología asociada |
|---|---|---|---|---|
| Tipo I y II | Queratinas ácidas y básicas (>50 tipos) | Células epiteliales (específico según epitelio) | Resistencia mecánica · anclaje a desmosomas y hemidesmosomas | Epidermólisis bullosa simple · ictiosis · tumores (marcador diagnóstico) |
| Tipo III | Vimentina | Células mesenquimales: fibroblastos · endotelio · linfocitos | Forma celular · migración · señalización mecanosensorial | Marcador de sarcomas y transición epitelio-mesénquima |
| Tipo III | Desmina | Músculo esquelético · cardíaco · liso | Conecta sarcómeros · ancla miofilamentos a membrana | Miopatías por desmina · cardiomiopatía dilatada |
| Tipo III | GFAP | Astrocitos del SNC | Soporte estructural de la glía | Marcador de gliomas · enfermedad de Alexander |
| Tipo IV | Neurofilamentos (NF-L, NF-M, NF-H) | Neuronas (especialmente axones) | Determinan calibre axonal · velocidad de conducción nerviosa | Acumulación en ELA · enfermedad de Charcot-Marie-Tooth |
| Tipo V | Laminas A, B y C | Núcleo de todas las células eucariotas | Lámina nuclear densa · anclaje de cromatina · estructura nuclear | Laminopatías (progeria · distrofia muscular de Emery-Dreifuss) |
Las laminas nucleares, su papel en la estructura del núcleo y las laminopatías se desarrollan con detalle en T13 — Arquitectura del núcleo interfásico, apartado 13.4.1.
La identificación del tipo de filamento intermedio expresado por las células tumorales es una herramienta diagnóstica fundamental en anatomía patológica para determinar el origen de tumores poco diferenciados:
- Queratinas positivas: tumor de origen epitelial (carcinoma).
- Vimentina positiva: tumor de origen mesenquimal (sarcoma).
- GFAP positiva: tumor glial (glioma, astrocitoma).
- Desmina positiva: tumor muscular (rabdomiosarcoma).
- Las epidermólisis bullosas simples son enfermedades genéticas causadas por mutaciones en las queratinas 5 y 14, que forman los filamentos intermedios de los queratinocitos basales. Sin queratinas funcionales, la capa basal de la epidermis es frágil y se rompe ante traumatismos mínimos, produciendo ampollas intraepidérmicas.
