11.1. Caracteristicas generales de la transcripcion
La transcripción es el proceso por el cual la información contenida en una secuencia de ADN se copia en forma de ARN. Se trata de un proceso:
- Conservativo: el ADN utilizado como molde permanece intacto en la célula.
- Selectivo: solo se transcribe una de las dos hebras del ADN (la hebra molde o antisentido) y solo los genes que la célula necesita en un momento dado. Esta selectividad convierte a la transcripción en el principal punto de control de la expresión génica.
- Independiente del ciclo celular: puede producirse en cualquier momento de la vida celular.
- Unidireccional: la síntesis del ARN se da en sentido 5′ a 3′.
A diferencia de la replicación, la ARN polimerasa no dispone de actividad correctora de errores, lo que resulta en una tasa de error de 10-5 a 10-6 (entre dos y tres órdenes de magnitud mayor que la de la ADN polimerasa). Esta tasa de error relativa se tolera porque cada gen se transcribe múltiples veces y cada ARNm tiene una vida media corta en la célula. La velocidad de síntesis es de aproximadamente 50 nucleótidos por segundo.
Transcripción: proceso de síntesis de ARN a partir de una hebra molde de ADN, catalizada por la ARN polimerasa.
Características: conservativa, selectiva, unidireccional (5 a 3), sin cebador, sin corrección de errores, independiente del ciclo celular. Principal nivel de control de la expresión génica.
11.2. ARN polimerasas: tipos en procariotas y eucariotas
Las ARN polimerasas catalizan la adición de ribonucleótidos trifosfato (NTPs) al extremo 3-OH del ARN creciente, con liberación de pirofosfato que es hidrolizado irreversiblemente, impulsando termodinámicamente la reacción. A diferencia de las ADN polimerasas, no necesitan cebador para iniciar la síntesis.
La ARN polimerasa procariótica (holoenzima) es un enzima complejo de 450 kDa formado por el núcleo enzimático (subunidades alfa-2, beta y beta-prima) responsable de la polimerización, más la subunidad sigma.
El núcleo realiza la catális de síntesis, separación de hebras, desplazamiento y desenrollamiento. La subunidad sigma (de la que existen distintos tipos en función del estrés celular) proporciona la capacidad de reconocer promotores específicos e iniciar la transcripción en el lugar correcto. Una vez iniciada la elongación, sigma se disocia.
Los eucariotas poseen tres ARN polimerasas nucleares especializadas.
- La ARN polimerasa I transcribe los genes del ARN ribosomal grande (28S, 18S, 5.8S), los de mayor demanda celular.
- La ARN polimerasa II transcribe los genes que codifican proteínas (produciendo los ARN heterogéneos nucleares que se procesan hasta ARNm) y la mayoría de los ARN no codificantes reguladores.
- La ARN polimerasa III transcribe genes de ARN pequeños, principalmente ARNt y el ARN ribosomal 5S.
Ninguna de las tres ARN polimerasas puede actuar directamente sobre el ADN sin el concurso de factores de transcripción generales.
La especialización de las ARN polimerasas eucarióticas permite regular por separado la síntesis de los distintos tipos de ARN. La ARN pol II es la de mayor relevancia en la regulación de la expresión génica, ya que transcribe los genes codificantes de proteínas. Su actividad se regula por fosforilación del dominio carboxilo-terminal (CTD).
11.3. Promotores y secuencias consenso
Promotor: secuencia de ADN que indica a la ARN polimerasa donde debe iniciar la transcripción, en qué condiciones y en qué sentido. Define el gen que se va a transcribir. En procariotas, la subunidad sigma reconoce directamente el promotor; en eucariotas, los factores de transcripción generales construyen el complejo de preiniciación sobre el promotor y reclutan a la ARN pol II.
El concepto de secuencia consenso es esencial para entender la lógica de los promotores. Una secuencia consenso no es una secuencia real, sino la secuencia ideal obtenida por extrapolación estadística tras analizar múltiples promotores: se asigna a cada posición el nucleótido más frecuente entre todos los promotores comparados. Cuanto más parecida sea la secuencia real de un promotor a la consenso, más fuerte y activo será ese promotor; las desviaciones de la consenso reducen la eficiencia de inicio. En procariotas, las regiones consenso más estudiadas (caja Pribnow, TATAAT) se sitúan a -10 y -35 respecto al sitio de inicio de la transcripción.
En eucariotas, los promotores de la ARN pol II presentan una arquitectura más compleja: contienen la caja TATA (reconocida por el factor TFIID), elementos próximos al promotor (GC box, CAAT box) y potenciadores (enhancers) que pueden estar a miles de nucleótidos de distancia y actúan aumentando la actividad transcripcional al contactar con el promotor por lazo del ADN.
11.4. Mecanismo de la transcripción: iniciación, elongación y terminación
11.4.1. Iniciación
La iniciación de la transcripción en procariotas sigue una secuencia bien definida.
- La holoenzima (núcleo + sigma) se une inicialmente al ADN de forma no especifica y difunde hasta que sigma reconoce el promotor. Se forma el complejo cerrado (promotor cerrado).
- A continuación, la ARN polimerasa funde localmente las hebras del ADN en la región alrededor del sitio de inicio, generando el complejo abierto (promotor abierto).
- Comienza la síntesis del ARN nucleótido a nucleótido. Durante los primeros 12 nucleótidos, el complejo es inestable y puede abortarse la transcripción
- Si se superan los 12 nucleótidos, la subunidad sigma se disocia y la ARN polimerasa sufre un cambio conformacional que estabiliza el complejo de elongación.
11.4.2. Elongación
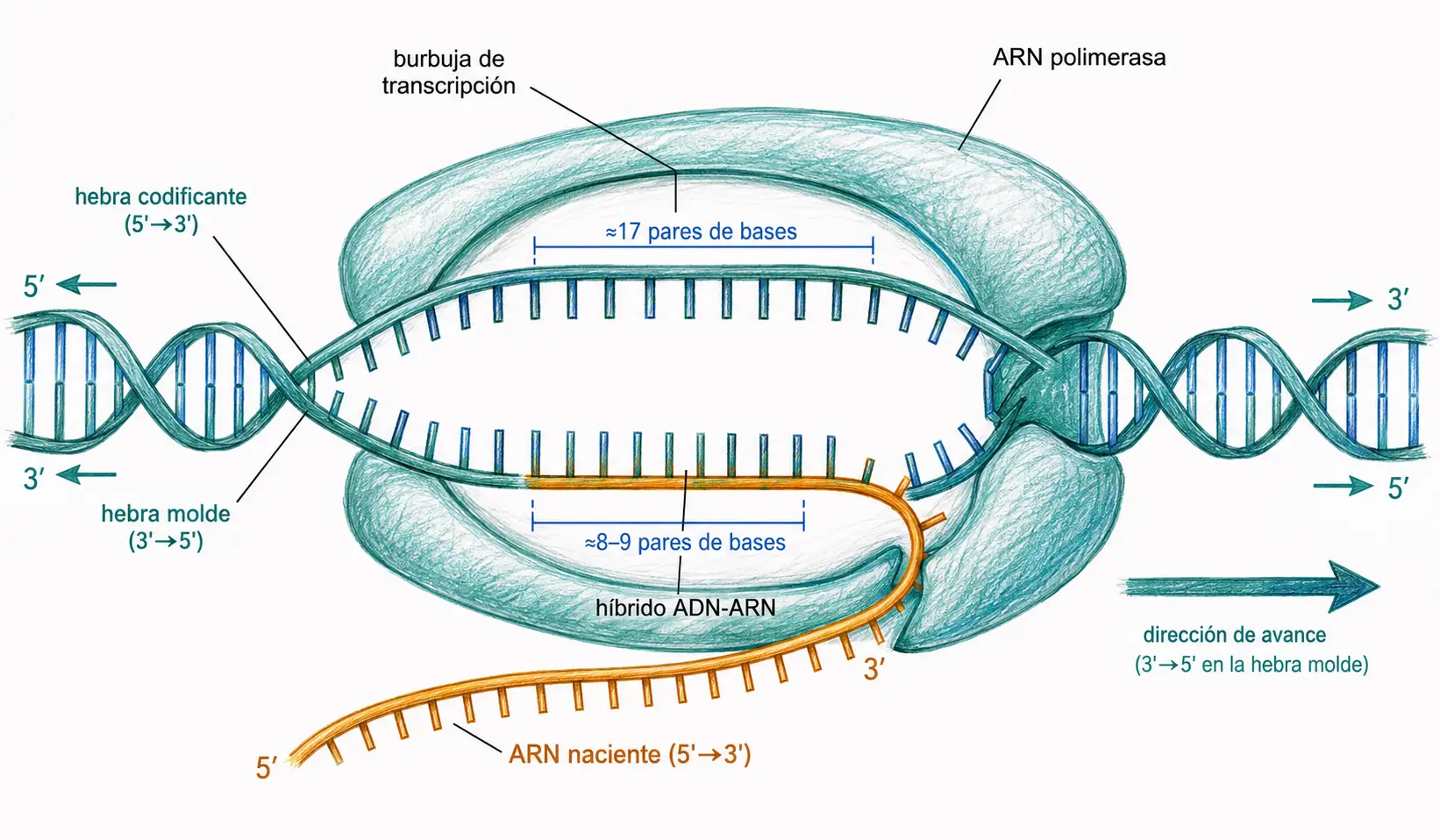
Durante la elongación, la ARN polimerasa genera una burbuja de transcripción de aproximadamente 17 pares de bases:
- El ADN bicatenario se mantiene abierto localmente mientras la polimerasa avanza sobre la hebra molde en dirección 3′ a 5′, sintetizando el ARN complementario en sentido 5′ a 3′.
- El ARN recién sintetizado se separa rápidamente del molde y el ADN se vuelve a cerrar en su estela.
11.4.3. Terminación
La terminación de la transcripción en procariotas puede ser dependiente o independiente (intrínseca) de proteínas.
- En la terminación dependiente de proteínas:
- La proteína rho (hexámero con actividad helicasa y ATPasa):
- Reconoce una secuencia específica del ARN.
- Asciende hacia la ARN polimerasa consumiendo ATP.
- Al alcanzarla desestabiliza el complejo y libera el transcrito.
- La proteína NusA (similar en función a la subunidad sigma) aumenta la sensibilidad de la ARN polimerasa a las señales de terminación internas.
- La proteína rho (hexámero con actividad helicasa y ATPasa):
- En la terminación intrínseca o independiente, la ARN polimerasa llega a una región del ADN que, al transcribirse, genera un ARN con dos características secuenciales:
- Primero una secuencia palindrómica rica en G-C que se pliega en horquilla, enlenteciendo la polimerasa.
- Inmediatamente después, una secuencia rica en A-T que proporciona pocos puentes de hidrogeno, de modo que el hibrido ADN-ARN es débil y la polimerasa se disocia.
Terminación intrínseca en procariotas: secuencia palindrómica en ARN (horquilla GC) que enlentece la ARN polimerasa, seguida de secuencia rica en AU (débil hibrido ADN-ARN) que permite la disociación.
Terminación dependiente de rho: proteína rho reconoce ARN, asciende con gasto de ATP, desestabiliza el complejo de transcripción.
11.5. Regulación de la transcripción
11.5.1. Regulación en procariotas: el operón
En procariotas, los genes que codifican enzimas de una misma ruta metabólica suelen estar agrupados en el cromosoma y se transcriben juntos bajo el control de un único promotor. Esta unidad funcional se denomina operón.
Un operón está formado por tres elementos: el gen regulador, que codifica una proteína reguladora (represor o activador); el centro de control, que incluye el promotor (donde se une la ARN polimerasa) y el operador (donde se une la proteína reguladora); y los genes estructurales, que codifican las proteínas cuya expresión se regula.
Operón: unidad de expresión génica procariótica formada por un gen regulador, un centro de control (promotor + operador) y un conjunto de genes estructurales transcritos como una unidad. Permite la expresión coordinada de genes funcionalmente relacionados.
La proteína reguladora puede actuar como represor (impide la transcripción al unirse al operador) o como activador (la facilita al contactar con la ARN polimerasa). En ambos casos, su actividad depende de la unión a un ligando de pequeña molécula que modifica su conformación y, por tanto, su afinidad por el ADN.
El operón lac: regulación negativa con inducción por ligando
El operón lac de Escherichia coli es el modelo canónico de operón catabólico. Codifica tres enzimas necesarias para metabolizar la lactosa como fuente de carbono, entre ellas la β-galactosidasa, que rompe el disacárido en glucosa y galactosa.
En ausencia de lactosa, el represor lac está unido al operador y bloquea el avance de la ARN polimerasa. Cuando la lactosa está presente, parte de ella se convierte intracelularmente en alolactosa, un isómero que actúa como inductor: se une al represor lac, provoca un cambio conformacional y reduce drásticamente su afinidad por el operador. El represor se disocia, el operador queda libre y la ARN polimerasa puede transcribir los genes estructurales.
El operón lac tiene además un nivel de regulación positiva: cuando la glucosa escasea, la enzima adenilato ciclasa produce AMPc. El complejo AMPc-CAP (proteína activadora de catabolitos) se une al promotor lac y aumenta la afinidad de la ARN polimerasa por ese promotor. Solo cuando hay lactosa Y no hay glucosa se obtiene la máxima transcripción del operón.
Esto tiene sentido energético: la bacteria solo invierte en sintetizar los enzimas de degradación de lactosa cuando la lactosa está disponible (señal del represor) y cuando la fuente de carbono preferida, la glucosa, no lo está (señal del AMPc-CAP).
El operón Trp: regulación negativa con represión por producto final
El operón Trp es un ejemplo de operón anabólico: sus cinco genes estructurales codifican enzimas que sintetizan triptófano a partir del precursor corismato. La lógica reguladora es la inversa al operón lac.
El represor TrpR está inactivo (no puede unirse al operador) mientras el nivel de triptófano es bajo. Cuando el triptófano se acumula, actúa como corepressor: se une al TrpR, cambia su conformación y activa su unión al operador, reprimiendo la transcripción de los genes de síntesis. La célula deja de fabricar lo que ya tiene en exceso.
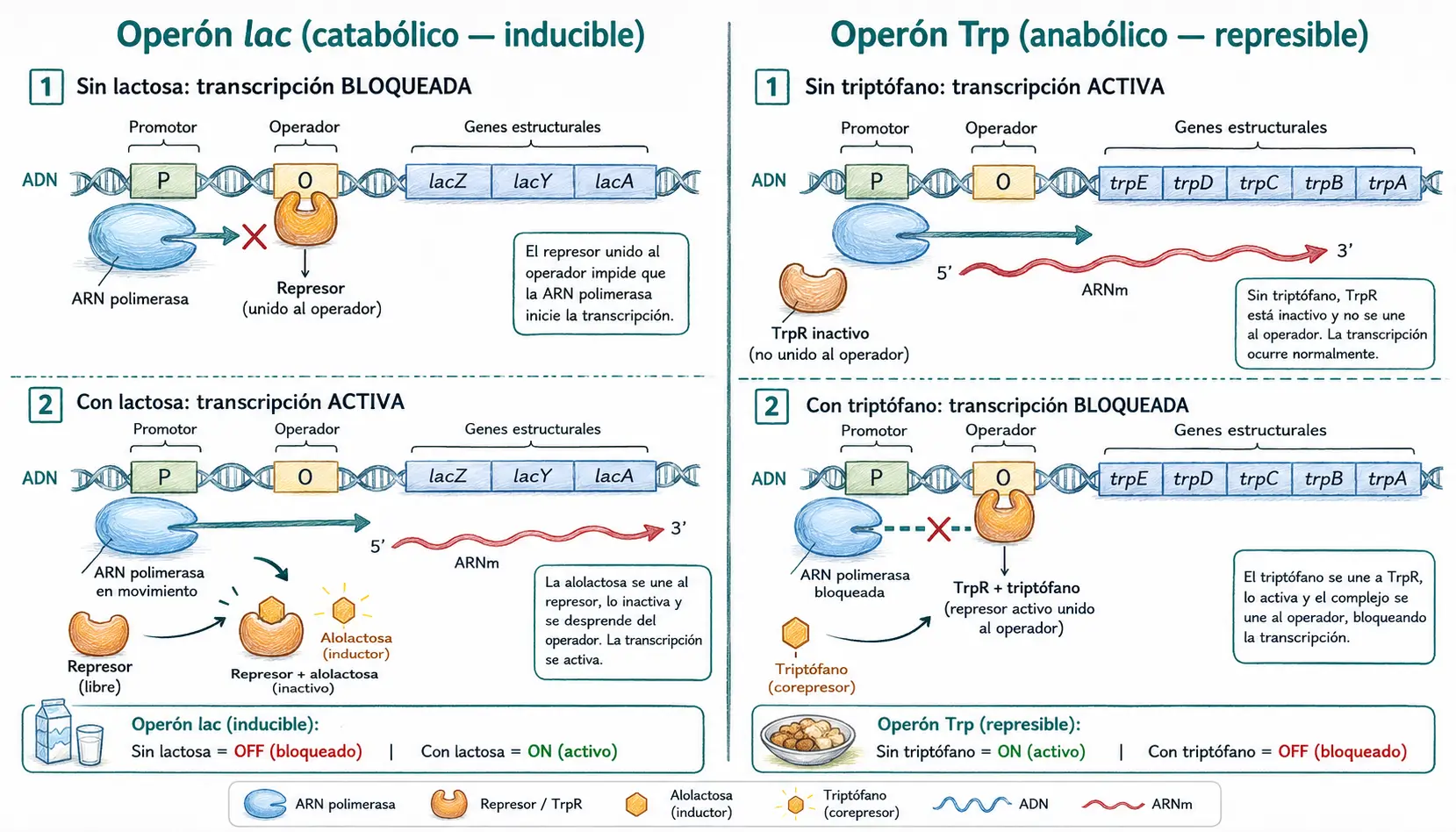
Diferencia operón lac vs. operón Trp:
En el operón lac (catabólico), el ligando (alolactosa) disocia el represor del ADN → se activa la transcripción.
En el operón Trp (anabólico), el ligando (triptófano) une el represor al ADN → se reprime la transcripción.
La lógica es siempre de ahorro: se transcribe cuando se necesita el producto, se reprime cuando se tiene de sobra.
En eucariotas no existen operones. Cada gen tiene su propio promotor y se regula de forma independiente. La regulación coordinada de genes relacionados se logra mediante factores de transcripción compartidos que reconocen elementos reguladores en múltiples promotores dispersos por el genoma. (→ ver apartado 11.5.2 de este mismo tema)
11.5.2. Factores de transcripción en eucariotas: motivos de unión al ADN y receptores nucleares
En eucariotas, la regulación de la transcripción recae sobre proteínas modulares denominadas factores de transcripción que reconocen secuencias específicas del ADN y modulan la actividad de la ARN polimerasa II. Estos factores tienen dos tipos de dominios: funcionalmente diferenciados:
- El dominio de unión al ADN.
- El dominio de activación o represión.
La modularidad es funcional: los dos dominios pueden separarse e incluso intercambiarse entre proteínas sin perder su función individual, lo que ha permitido una enorme diversificación evolutiva.
Los factores de transcripción no actúan solos sobre el promotor mínimo. Se unen a elementos reguladores proximales (a menos de 200 pb del sitio de inicio) y a potenciadores o enhancers (secuencias de 100–200 pb que pueden estar a 50 kb de distancia o más, en cualquier orientación y en intrones o regiones 3′). Los enhancers contactan con el promotor por plegamiento del ADN, formando un lazo que acerca factores distantes. La unión cooperativa de múltiples activadores en sitios próximos genera un complejo multiproteico denominado amplificosoma.
Motivos estructurales de unión al ADN
Los dominios de unión al ADN presentan estructuras recurrentes bien caracterizadas.
- El motivo hélice-bucle-hélice (HLH) consta de dos hélices alfa unidas por un bucle flexible. Una de las hélices (la de reconocimiento) entra en el surco mayor del ADN y establece contactos específicos con las bases; la otra estabiliza la estructura por interacciones hidrofóbicas con la primera. Estas proteínas actúan siempre como dímeros, lo que duplica la superficie de contacto con el ADN y aumenta la especificidad.
- El homeodominio se pliega en tres hélices alfa empaquetadas mediante interacciones hidrofóbicas. Las hélices 2 y 3 forman un motivo hélice-giro-hélice similar al HLH, con la hélice 3 interaccionando con el surco mayor. Los homeodominios son frecuentes en factores de transcripción que controlan el desarrollo embrionario.
- El motivo cremallera de leucina combina un dominio básico N-terminal (que contacta con el ADN) y un dominio C-terminal con leucinas cada siete posiciones que forman una interfase de dimerización hidrofóbica. Los dímeros resultantes se unen al ADN como una pinza sobre dos surcos mayores adyacentes.
- Los dedos de zinc son dominios en los que la cadena polipeptídica se pliega alrededor de uno o dos iones Zn²⁺. Existen varias clases según los ligandos del zinc: los dedos C2H2 (dos cisteínas y dos histidinas, monoméricos) son los más abundantes en el genoma humano y se unen en tándem al ADN; los C4 (cuatro cisteínas, diméricos) caracterizan a los receptores nucleares de la superfamilia de receptores de esteroides.
| Motivo | Estructura | Dimerización | Surco de unión | Ejemplo |
|---|---|---|---|---|
| HLH Hélice-bucle-hélice | 2 hélices α unidas por bucle flexible | Obligatoria (homodímero o heterodímero) | Surco mayor | MyoD, factores bHLH |
| HD Homeodominio | 3 hélices α empaquetadas; hélice 3 de reconocimiento | No obligatoria | Surco mayor | Factores Hox, Pax |
| bZIP Cremallera de leucina | Dominio básico N-t + Leu cada 7 posiciones C-t | Obligatoria (pinza sobre 2 surcos mayores) | Surco mayor | c-Jun, c-Fos, CREB |
| ZF Dedo de zinc C2H2 | Bucle sobre Zn²⁺ (2 Cys + 2 His) | Monomérico (múltiples dedos en tándem) | Surco mayor | SP1, TFIIIA |
| NR Dedo de zinc C4 (receptor nuclear) | Bucle sobre Zn²⁺ (4 Cys) | Obligatoria (homodímero o heterodímero) | Surco mayor | GR, RAR, TR, VDR |
Los cuatro motivos de unión al ADN más frecuentes son: hélice-bucle-hélice (HLH), homeodominio, cremallera de leucina y dedos de zinc. Todos interaccionan con el ADN a través del surco mayor. Los HLH y las cremalleras de leucina actúan obligatoriamente como dímeros.
Receptores nucleares y hormonas lipofílicas
Una clase especialmente relevante de factores de transcripción son los receptores nucleares, que pertenecen a la superfamilia de factores con dedo de zinc C4. Actúan como factores de transcripción dependientes de ligando: en ausencia del ligando suelen estar en el citoplasma o inactivos en el núcleo; al unirse al ligando cambian de conformación, se translocan al núcleo (si no estaban ya allí), dimerizan y se unen al ADN.
Sus ligandos son hormonas lipofílicas que atraviesan la membrana plasmática por difusión: hormonas esteroideas (cortisol, estrógenos, testosterona), hormona tiroidea (T3/T4) y vitaminas liposolubles con función hormonal (ácido retinoico, vitamina D).
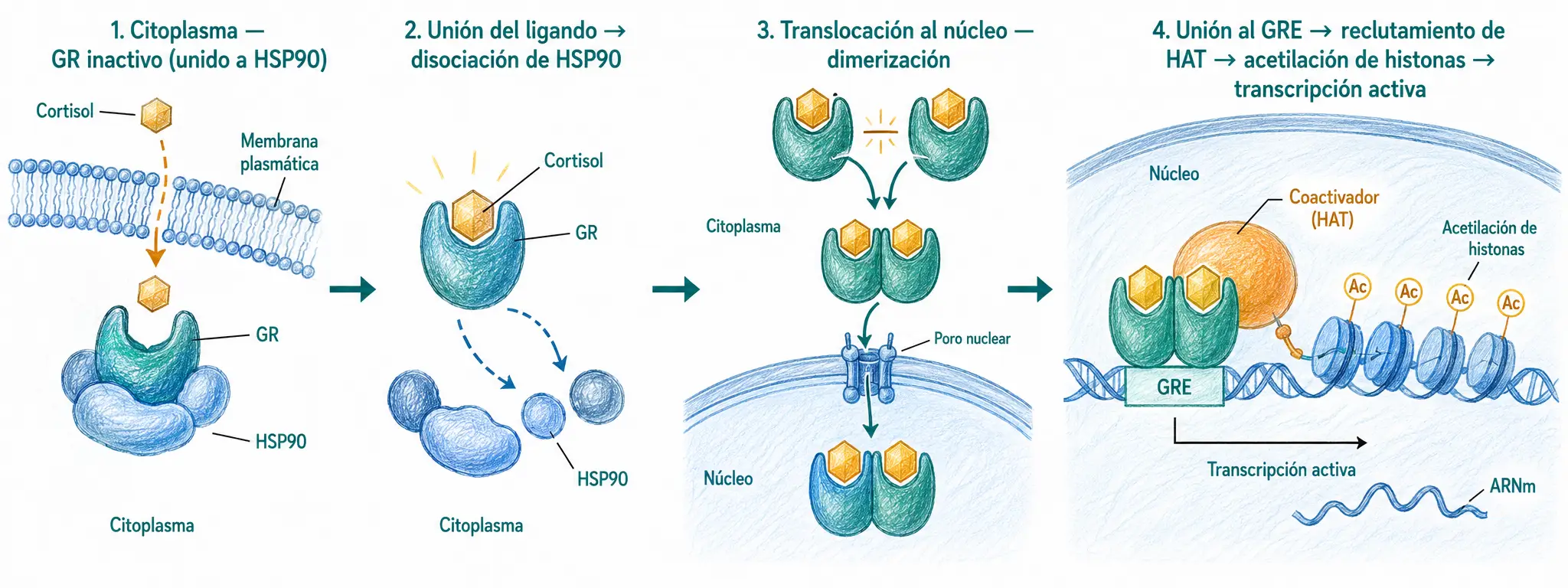
El receptor de glucocorticoides (GR) es el ejemplo canónico. En ausencia de cortisol permanece en el citoplasma asociado a proteínas chaperonas (HSP90). Cuando el cortisol entra en la célula y se une al GR, las chaperonas se disocian, el complejo se transloca al núcleo, dimeriza y se une a elementos de respuesta a glucocorticoides (GRE) en el ADN, activando genes antiinflamatorios e inhibiendo la transcripción de genes proinflamatorios. Este mecanismo explica el potente efecto antiinflamatorio de los corticoides exógenos.
Remodelación de cromatina y acetilación de histonas
La accesibilidad del promotor al complejo de preiniciación depende del estado de compactación de la cromatina. En la cromatina condensada (heterocromatina), el ADN está enrollado estrechamente sobre los nucleosomas y los promotores son inaccesibles. Para que la transcripción se active, los nucleosomas deben desplazarse.
Cuando un activador se une a su elemento regulador, recluta coactivadores con actividad histona acetiltransferasa (HAT). Estas enzimas transfieren grupos acetilo a los residuos de lisina de las colas N-terminales de las histonas. La acetilación neutraliza la carga positiva de las lisinas, reduciendo su afinidad por el ADN cargado negativamente, lo que abre la cromatina localmente. Además, las lisinas acetiladas son reconocidas por complejos remodeladores de cromatina dependientes de ATP (como SWI/SNF) que utilizan energía para desplazar físicamente los nucleosomas y dejar el promotor expuesto.
El proceso es reversible: las histona desacetilasas (HDAC) eliminan los grupos acetilo y restauran la compactación de la cromatina, silenciando la transcripción. El equilibrio HAT/HDAC sobre un promotor determinado es uno de los principales mecanismos de regulación epigenética de la expresión génica.
Las modificaciones de histonas (acetilación, metilación, fosforilación) forman parte del código de histonas y constituyen un nivel de regulación epigenética que se estudia en detalle en el Tema 14 - Niveles post-traduccionales de control de la expresión génica).
11.6. Comparación entre replicación y transcripción
Aunque replicación y transcripción comparten el uso del ADN como molde y la síntesis en sentido 5′ a 3′, presentan diferencias fundamentales:
- La replicación es semiconservativa (cada hebra nueva se aparea con una parental) y produce ADN; la transcripción es conservativa (el ADN molde permanece intacto) y produce ARN.
- La replicación es total (se copia todo el genoma) y esta ligada al ciclo celular; la transcripción es selectiva y continua.
- La ADN polimerasa requiere cebador y tiene actividad correctora de errores (tasa de error 10-7 a 10-8, velocidad 1000 nt/s); la ARN polimerasa no requiere cebador, no corrige errores (tasa 10-5 a 10-6, velocidad 50 nt/s).
- La replicación garantiza la operatividad y fidelidad del genoma; la transcripción controla la expresión génica.
| Característica | Replicación | Transcripción |
|---|---|---|
| Producto | ADN (doble hélice hija) | ARN (copia de un gen) |
| Mecanismo | Semiconservativo | Conservativo |
| Alcance | Total (todo el genoma) | Selectivo (genes necesarios) |
| Relación con ciclo celular | Ligada (fase S) | Independiente |
| Direccionalidad | Bidireccional (5'→3' en cada hebra) | Unidireccional (5'→3') |
| Cebador | Sí (ARN cebador obligatorio) | No (inicio de novo) |
| Corrección de errores | Sí (exonucleasa 3'→5') | No |
| Tasa de error | 10⁻⁸ – 10⁻⁹ | 10⁻⁵ – 10⁻⁶ |
| Velocidad | 1000 nt/s (procariotas) | 50 nt/s |
| Enzima principal | ADN polimerasa | ARN polimerasa |
| Función biológica | Fidelidad y transmisión del genoma | Control de la expresión génica |
La mayor tasa de error de la ARN polimerasa frente a la ADN polimerasa es biologicamente aceptable porque:
- Cada gen se transcribe en múltiples copias.
- Los ARNm tienen vida media corta y se reemplazan.
- Los errores no se heredan. En cambio, los errores de replicacion si pueden heredarse, de ahi la necesidad de correccion de pruebas.
