13.1. El código genético
La traducción necesita una clave que asigne a cada combinación de nucleótidos del ARNm un aminoácido concreto. Esa clave es el código genético: el conjunto de reglas que relacionan la secuencia de nucleótidos del ARNm con la secuencia de aminoácidos de la proteína.
La unidad fundamental del código es el codón o triplete: una secuencia de tres nucleótidos consecutivos en el ARNm. Con cuatro bases posibles en cada posición, existen 4³ = 64 combinaciones posibles. Como solo hay 20 aminoácidos estándar, el código tiene más codones de los estrictamente necesarios.
Código genético: sistema de correspondencia entre tripletes de nucleótidos (codones) del ARNm y aminoácidos. Consta de 64 codones: 61 codifican aminoácidos y 3 son codones de terminación (UAA, UAG, UGA). El codón de inicio es AUG (metionina).
El código genético tiene cinco propiedades fundamentales:
- Es degenerado: la mayoría de los aminoácidos están codificados por más de un codón. Solo la metionina (AUG) y el triptófano (UGG) tienen un único codón. Los codones que codifican el mismo aminoácido se denominan codones sinónimos y normalmente comparten las dos primeras bases.
- Es no ambiguo: un codón dado siempre especifica el mismo aminoácido. Un triplete no puede codificar dos aminoácidos distintos.
- Es secuencial y sin solapamiento: los codones se leen de forma consecutiva, en grupos de tres, sin que los nucleótidos de un codón formen parte del siguiente. La pauta de lectura (reading frame) queda fijada desde el codón AUG de inicio.
- Es universal con pequeñas excepciones: el mismo código rige en prácticamente todos los organismos conocidos, desde bacterias hasta humanos. Las excepciones se encuentran en genomas de orgánulos (mitocondrias, cloroplastos) y en algunos microorganismos, pero siempre son cambios menores.
- Es continuo: una vez iniciada la traducción en AUG, los codones se leen ininterrumpidamente hasta que un codón de terminación entra en el sitio A del ribosoma.
La degeneración del código se concentra en la tercera base del codón (posición 3′ o posición de balanceo, wobble position). El apareamiento entre la tercera base del codón y la primera base del anticodón es más flexible estéricamente que el de las dos primeras posiciones, lo que permite a un único ARNt reconocer varios codones sinónimos que difieren solo en la tercera base.
Existe un segundo código genético implícito: la correspondencia entre cada ARNt y su aminoácido correcto, establecida por las aminoacil-ARNt sintetasas. Este segundo código determina la fidelidad real de la traducción y se explica en el apartado siguiente.
13.2. ARNt y aminoacil-ARNt sintetasas
13.2.1. Estructura y función del ARNt
Los ARN de transferencia (ARNt) son los adaptadores moleculares que conectan el lenguaje de los nucleótidos con el de los aminoácidos. Tienen una longitud de 73–93 nucleótidos y se pliegan en una estructura secundaria en forma de trébol de cuatro brazos, que a su vez adopta una conformación tridimensional en forma de L.
Dos regiones del ARNt son funcionalmente importantes:
- El bucle del anticodón, situado en el extremo opuesto de la molécula, contiene la secuencia de tres nucleótidos que se aparean por complementariedad con el codón del ARNm.
- El extremo 3′ CCA, invariante en todos los ARNt, al que se une covalentemente el aminoácido mediante un enlace éster.
Existen entre 40 y 60 especies distintas de ARNt en la célula, suficientes para reconocer los 61 codones sentido gracias al balanceo de la tercera base.
13.2.2. Aminoacil-ARNt sintetasas: el segundo código genético
Las aminoacil-ARNt sintetasas (aaRS) son las enzimas responsables de cargar cada ARNt con su aminoácido correcto. Existen 20 tipos, una por aminoácido. La reacción que catalizan es la activación del aminoácido, que ocurre en dos pasos:
- El aminoácido reacciona con ATP formando un intermediario aminoacil-adenilato (aa~AMP) con liberación de pirofosfato (PPi).
- El aminoácido se transfiere desde el intermediario al extremo 3′-OH del ARNt correspondiente, liberando AMP. El PPi es hidrolizado a 2 Pi por pirofosfatasas celulares, haciendo la reacción global irreversible.
La activación de un aminoácido consume el equivalente a dos enlaces de alta energía del ATP: se hidroliza ATP → AMP + PPi, y el PPi se hidroliza a 2 Pi. El enlace éster aminoacil-ARNt resultante es un enlace activado: la energía almacenada en él se utilizará para formar el enlace peptídico en el ribosoma.
La fidelidad de las aminoacil-ARNt sintetasas es crítica para la precisión de toda la traducción. El ribosoma lee el anticodón del ARNt, no el aminoácido; si una sintetasa carga un aminoácido incorrecto en un ARNt, ese error se propagará al polipéptido sin posibilidad de corrección posterior. Por eso estas enzimas disponen de un sitio de corrección (editing site): si el aminoácido unido al ARNt es incorrecto, la enzima hidroliza el enlace éster y repite la reacción.
La fidelidad de la traducción no la garantiza el ribosoma: la garantizan las aminoacil-ARNt sintetasas. El ribosoma comprueba que el anticodón del ARNt se aparea correctamente con el codón del ARNm, pero no puede verificar qué aminoácido lleva ese ARNt.
13.3. El ribosoma
El ribosoma es la maquinaria molecular donde se produce la síntesis del enlace peptídico. Es un complejo ribonucleoproteico formado por dos subunidades de tamaños desiguales que se ensamblan sobre el ARNm durante la iniciación y se disocian al final de la traducción.
- En procariotas, el ribosoma completo (monosoma) sedimenta a 70S y está formado por la subunidad pequeña 30S (ARNr 16S + ~21 proteínas) y la subunidad grande 50S (ARNr 23S + ARNr 5S + ~31 proteínas).
- En eucariotas, el ribosoma completo sedimenta a 80S y está formado por la subunidad pequeña 40S (ARNr 18S + ~33 proteínas) y la subunidad grande 60S (ARNr 28S + ARNr 5.8S + ARNr 5S + ~49 proteínas).
Ribosoma procariota: 70S = 30S + 50S. Ribosoma eucariota: 80S = 40S + 60S. Los valores S (Svedberg) no son aditivos porque dependen de la forma, no solo del tamaño: 30S + 50S = 70S, no 80S.
El ribosoma ensamblado presenta tres sitios funcionales en la interfase entre las dos subunidades:
- El sitio A (aminoacil) es el punto de entrada del aminoacil-ARNt, el que porta el nuevo aminoácido que se va a incorporar.
- El sitio P (peptidil) sostiene el ARNt unido a la cadena polipeptídica naciente.
- El sitio E (exit) retiene el ARNt desacilado (ya sin aminoácido) justo antes de que abandone el ribosoma.
La actividad peptidil transferasa, responsable de catalizar la formación del enlace peptídico, reside en el ARNr 23S (procariotas) o 28S (eucariotas) de la subunidad mayor. Es una ribozima: las proteínas ribosomales tienen un papel estructural, pero la catálisis química la realiza el ARN.
| Característica | Procariotas | Eucariotas |
|---|---|---|
| Ribosoma completo | 70S | 80S |
| Subunidad pequeña | 30S (ARNr 16S) | 40S (ARNr 18S) |
| Subunidad grande | 50S (ARNr 23S + 5S) | 60S (ARNr 28S + 5.8S + 5S) |
| Reconocimiento del inicio | Secuencia Shine-Dalgarno / ARNr 16S | Cap m7G / scanning hasta primer AUG (Kozak) |
| Primer aminoácido | Formilmetionina (fMet) | Metionina (Met) |
| ARNm | Policistrónico | Monocistrónico |
| Peptidil transferasa (ribozima) | ARNr 23S | ARNr 28S |
| Diana de antibióticos | Sí (estreptomicina, eritromicina, tetraciclina…) | No (selectividad terapéutica) |
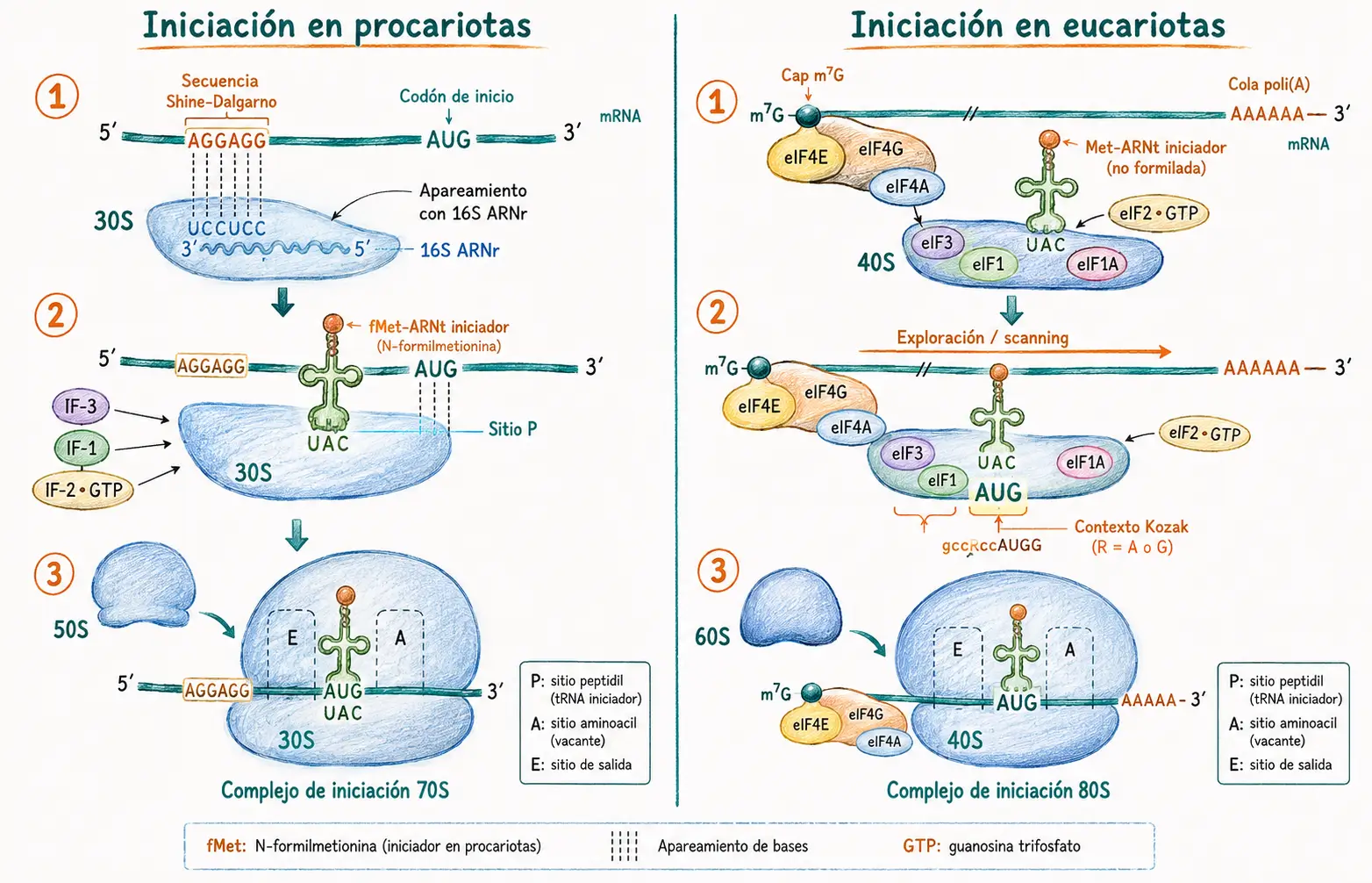
13.4. Iniciación de la traducción
La iniciación es la fase en que el ribosoma se ensambla sobre el ARNm en el codón de inicio AUG y el primer aminoacil-ARNt ocupa el sitio P.
13.4.1. Iniciación en procariotas
En procariotas, el reconocimiento del codón AUG de inicio depende de la secuencia de Shine-Dalgarno: una secuencia corta rica en purinas situada entre 5 y 10 nucleótidos antes del AUG en el ARNm, cuya secuencia es complementaria a una región del extremo 3′ del ARNr 16S de la subunidad 30S. Este apareamiento posiciona la subunidad pequeña directamente sobre el codón de inicio correcto.
El ensamblaje del complejo de iniciación 30S requiere los factores de iniciación IF-1 e IF-3 (que mantienen las subunidades separadas e impiden la unión prematura de la subunidad 50S), el ARNm, y el ARNt iniciador cargado con formilmetionina (fMet-ARNt^fMet), transportado por IF-2 en presencia de GTP. Una vez formado este complejo, la subunidad 50S se une, los factores IF se disocian y el GTP se hidroliza, completando el complejo de iniciación 70S con la fMet-ARNt^fMet en el sitio P.
En procariotas, un mismo ARNm puede contener varias secuencias Shine-Dalgarno precediendo distintos AUG, lo que permite la traducción de varias proteínas distintas a partir de un único transcrito (ARNm policistrónico). En eucariotas esto no ocurre: los ARNm son monocistónicos.
13.4.2. Iniciación en eucariotas
En eucariotas, el ribosoma 40S no busca secuencias complementarias en el ARNm. En cambio, la subunidad pequeña reconoce el cap m7G del extremo 5′ a través del factor eIF4E, que forma parte del complejo eIF4F. A continuación, el ribosoma cargado con el ARNt iniciador (Met-ARNt^Met, no formilado) explora el ARNm en dirección 5′→3′ hasta encontrar el primer AUG en contexto favorable (contexto Kozak: GCC(A/G)CCAUGG). Allí se une la subunidad 60S, se disocian los factores eIF y comienza la elongación.
La diferencia clave en la iniciación: los procariotas usan la secuencia Shine-Dalgarno para posicionarse directamente sobre el AUG correcto; los eucariotas usan el cap 5′ como punto de entrada y el modelo de exploración (scanning) para localizar el primer AUG.
13.5. Elongación
La elongación es el proceso cíclico por el cual se añaden sucesivamente aminoácidos a la cadena polipeptídica naciente. Cada ciclo incorpora un aminoácido y consta de tres pasos: entrada del aminoacil-ARNt, formación del enlace peptídico y translocación.
Paso 1 — Entrada del aminoacil-ARNt en el sitio A. El aminoacil-ARNt correcto (el cuyo anticodón es complementario al codón presente en el sitio A) llega al ribosoma transportado por el factor de elongación EF-Tu (en procariotas) cargado con GTP. Cuando el anticodón encaja correctamente con el codón, el GTP se hidroliza a GDP, el EF-Tu-GDP se disocia y el aminoacil-ARNt queda alojado en el sitio A. El tiempo que tarda la hidrólisis del GTP en producirse actúa como ventana de corrección: si el apareamiento anticodón-codón es incorrecto, el ARNt se disocia antes de que el GTP se hidrolice.
Paso 2 — Formación del enlace peptídico. La peptidil transferasa cataliza la transferencia de la cadena polipeptídica del ARNt del sitio P al aminoácido del ARNt del sitio A. Se forma un nuevo enlace peptídico y la cadena queda ahora unida al ARNt del sitio A, con un residuo más. El ARNt del sitio P queda desacilado.
Paso 3 — Translocación. El factor de elongación EF-G (translocasa), con hidrólisis de un GTP, desplaza el ribosoma tres nucleótidos en dirección 3′ sobre el ARNm. El ARNt-péptido pasa del sitio A al sitio P, el ARNt desacilado pasa del sitio P al sitio E y abandona el ribosoma, y el sitio A queda libre y expuesto al siguiente codón.
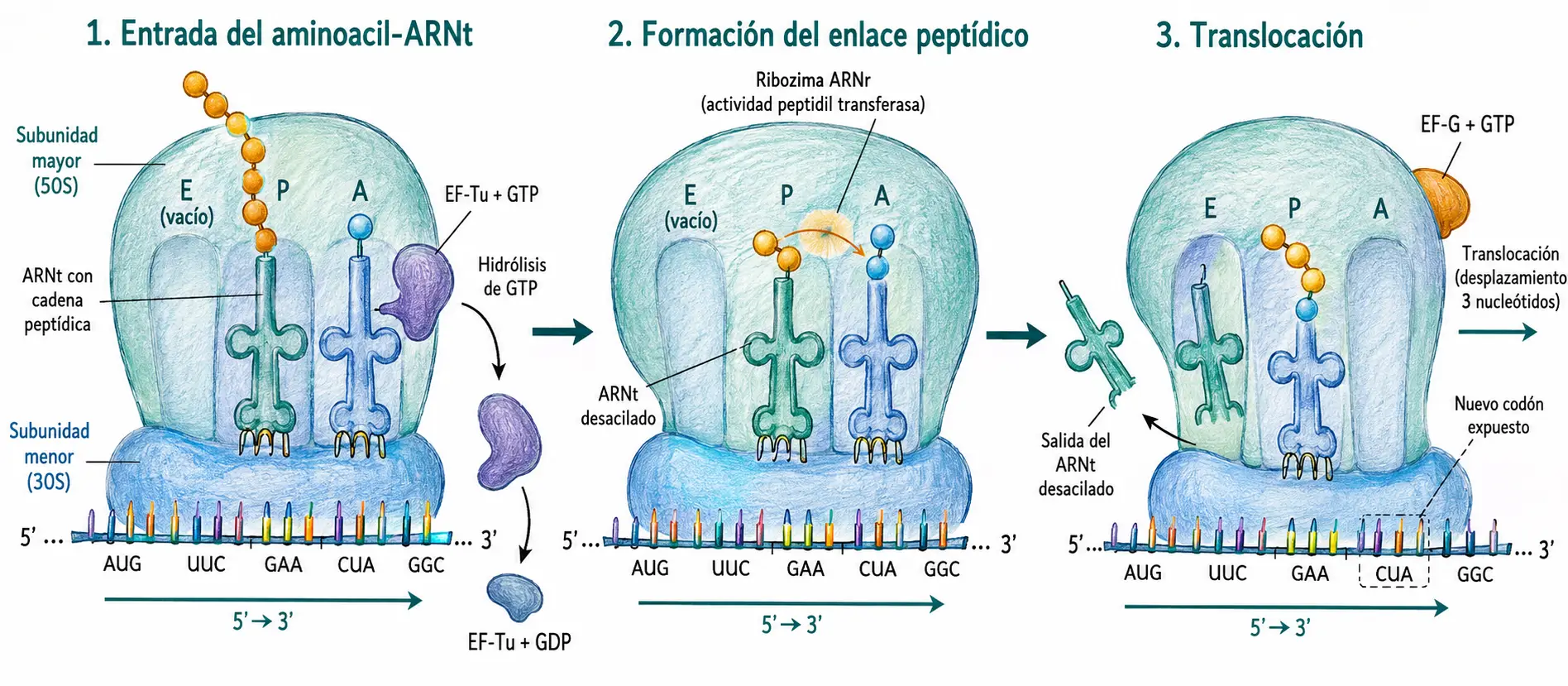
El coste energético de la elongación por ciclo es de 2 GTP (uno en la entrada del aminoacil-ARNt y uno en la translocación), más los 2 ATP equivalentes consumidos en la activación del aminoácido. Total: 4 enlaces ricos en energía por aminoácido incorporado.
El factor EF-Ts regenera el EF-Tu activo (EF-Tu-GTP) a partir del EF-Tu-GDP liberado tras la entrada del aminoacil-ARNt, cerrando el ciclo del factor de elongación.
13.6. Terminación
La elongación continúa hasta que uno de los tres codones de parada (UAA, UAG o UGA) entra en el sitio A del ribosoma. Ningún ARNt reconoce estos codones en condiciones normales. En su lugar, son reconocidos por factores de liberación (RF):
En procariotas, RF-1 reconoce UAA y UAG; RF-2 reconoce UAA y UGA; RF-3 es un factor auxiliar con actividad GTPasa que estimula la disociación de RF-1 y RF-2 tras la liberación del polipéptido.
Los factores de liberación mimetizan estructuralmente la forma de un ARNt y, al ocupar el sitio A, inducen un cambio conformacional en la peptidil transferasa que la convierte en una hidrolasa: en lugar de formar un enlace peptídico, cataliza la hidrólisis del enlace éster entre la cadena polipeptídica y el ARNt del sitio P. La cadena polipeptídica se libera, el ARNm se separa del ribosoma y las subunidades 30S y 50S se disocian, asistidas por los factores IF-1 e IF-3, quedando disponibles para un nuevo ciclo de traducción.
Algunos antibióticos actúan inhibiendo fases específicas de la traducción en ribosomas procariotas. La estreptomicina y los aminoglucósidos se unen a la subunidad 30S e interfieren con la corrección del apareamiento en el sitio A, causando errores de lectura. La eritromicina y los macrólidos bloquean la translocación uniéndose a la subunidad 50S. La tetraciclina impide la entrada del aminoacil-ARNt al sitio A. La selectividad por ribosomas 70S (procariotas) frente a 80S (eucariotas) es la base de su uso terapéutico.
13.7. Polisomas y eficiencia traductora
Una vez que el ribosoma iniciador ha avanzado suficientes nucleótidos desde el codón de inicio, el cap queda de nuevo accesible y puede ensamblarse un segundo ribosoma. Este proceso se repite, generando cadenas de ribosomas traduciendo simultáneamente el mismo ARNm denominadas polisomas (o polirribosomas).
Los polisomas aumentan la eficiencia traductora: a partir de un único ARNm se sintetizan múltiples copias de la proteína en paralelo. La densidad de ribosomas sobre un ARNm es proporcional a la tasa de iniciación y a la longitud del transcrito.
En procariotas, donde transcripción y traducción ocurren en el mismo compartimento y están acopladas temporalmente, los ribosomas pueden comenzar a traducir el extremo 5′ del ARNm mientras la ARN polimerasa todavía está sintetizando el extremo 3′. Este acoplamiento no existe en eucariotas, donde la transcripción ocurre en el núcleo y la traducción en el citoplasma.
