12.1. Introducción: del pre-ARNm al ARNm maduro
En eucariotas, la transcripción produce un ARN precursor largo e inmaduro denominado pre-ARNm o ARN heterogéneo nuclear (hnARN). Este transcrito primario no puede utilizarse directamente como molde para la síntesis de proteínas: contiene intrones intercalados entre los exones, sus extremos son vulnerables a la degradación y todavía se encuentra en el núcleo, separado de los ribosomas.
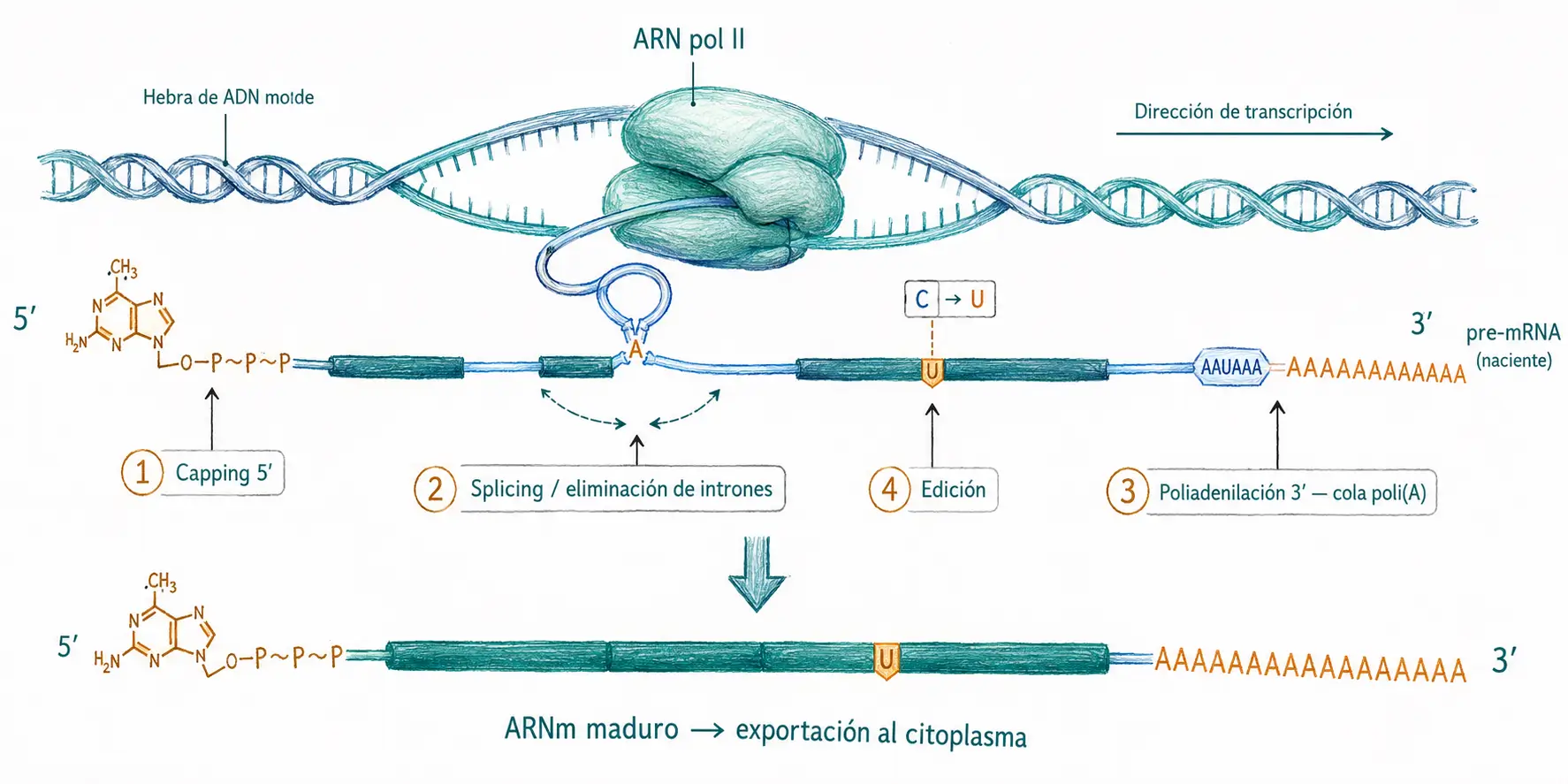
Para convertirse en un ARNm maduro y funcional, el pre-ARNm debe sufrir un conjunto de modificaciones covalentes en el núcleo. Estas modificaciones son cuatro:
- La adición de un cap en el extremo 5′.
- La poliadenilación del extremo 3′.
- El splicing (eliminación de intrones y unión de exones).
- En algunos transcritos, la edición de nucleótidos específicos.
No se producen de forma secuencial estricta: ocurren de manera cotranscripcional, es decir, mientras la ARN polimerasa II todavía está sintetizando el transcrito.
El nexo entre transcripción y procesamiento es el dominio carboxilo-terminal (CTD) de la ARN polimerasa II. Su hiperfosforilación durante la elongación lo convierte en una plataforma que recluta secuencialmente las enzimas de capping, los factores de splicing y la maquinaria de poliadenilación. Transcripción y procesamiento están físicamente acoplados.
Una vez completadas las modificaciones, el ARNm maduro se exporta al citoplasma a través de los poros nucleares. Allí queda disponible para los ribosomas, pero su destino final también depende de su estabilidad y de la acción de reguladores como los miARN. El conjunto de estos mecanismos determina cuánta proteína se sintetiza a partir de cada gen.
La transcripción y el papel del CTD fosforilado se explican con más detalle en T11: Síntesis del ARN (transcripción).
12.2. Adición del cap 5'
12.2.1. Capping: reacción y enzimas
Cuando el pre-ARNm emergente alcanza unos 25–30 nucleótidos de longitud, el CTD fosforilado recluta la maquinaria de capping. La reacción transcurre en tres pasos enzimáticos sobre el extremo 5′ del transcrito naciente.
El extremo 5′ de cualquier ARN recién sintetizado termina en un trifosfato (5′-ppp):
- Una ARN trifosfatasa elimina el fosfato terminal, dejando un extremo 5′-difosfato (5′-pp).
- Una guanililtransferasa añade una guanosina monofosfato (GMP) en orientación 5′→5′, formando el enlace trifosfato 5′–5′ característico del cap.
- Una ARN guanina-7-metiltransferasa transfiere un grupo metilo del donador SAM a la posición N7 de esa guanosina, generando la 7-metilguanosina (m7G).
El resultado es la estructura m7GpppN, donde N es el primer nucleótido del transcrito original. Esta es la forma básica denominada cap 0. En vertebrados, los nucleótidos adyacentes reciben metilaciones adicionales en la posición 2′-O de la ribosa (cap 1 y cap 2), lo que permite a la célula distinguir ARN propio de ARN viral.
12.2.2. Funciones del cap
El cap cumple tres funciones principales.
- Protege frente a la degradación: el enlace 5′–5′ trifosfato no puede ser atacado por las exonucleasas 5′→3′ normales.
- Facilita la exportación nuclear: las proteínas del complejo CBC (cap-binding complex) reconocen el cap en el núcleo y participan en el transporte del ARNm maduro a través del poro nuclear.
- Inicia la traducción: en el citoplasma, el factor eIF4E reconoce específicamente el cap m7G y recluta la maquinaria de iniciación ribosomal. Sin cap funcional, el ribosoma no puede unirse eficientemente al extremo 5′.
Muchos virus ARN utilizan estrategias para secuestrar eIF4E o sintetizar sus propios caps, compitiendo así con los ARNm celulares por la maquinaria traduccional. Esta dependencia del cap es el fundamento de varios mecanismos antivirales.
12.3. Poliadenilación del extremo 3'
12.3.1. Señal de poliadenilación y maquinaria molecular
La mayor parte de los ARNm eucariotas terminan en una larga cadena de adenosinas, la cola poli(A), que no está codificada en el ADN sino que se añade postranscripcionalmente. La señal de poliadenilación canónica es la secuencia AAUAAA, situada entre 10 y 30 nucleótidos por encima del sitio de corte.
El proceso es el siguiente:
- El complejo CPSF (cleavage and polyadenylation specificity factor) reconoce la secuencia AAUAAA.
- Otros factores proteicos (CstF, CFI, CFII) se unen a elementos cercanos y estabilizan el complejo.
- La endonucleasa del complejo corta el pre-ARNm en el sitio de poliadenilación, típicamente después de una CA.
- La ARN polimerasa II continúa sintetizando hacia abajo, pero ese fragmento se degrada por falta de cap.
Inmediatamente tras el corte, la poli(A) polimerasa (PAP) añade unos 200–250 residuos de adenosina al nuevo extremo 3′-OH sin necesidad de molde.
La cola poli(A) no la codifica el ADN ni la sintetiza la ARN polimerasa. La añade la poli(A) polimerasa después de que CPSF corta el pre-ARNm en el sitio de poliadenilación.
12.3.2. Funciones de la cola poli(A)
La cola poli(A) tiene dos funciones principales, operando sobre el extremo 3′.
- Protege frente a exonucleasas 3’→5′: actúa como tampón que retrasa la degradación desde el extremo 3′. Cuando la cola se acorta por debajo de cierto umbral, el ARNm queda desprotegido y se degrada rápidamente.
- Estimula la traducción: proteínas PABP (poly(A)-binding protein) citoplásmicas se unen a la cola poli(A) y contactan con eIF4G, que a su vez está unido al cap a través de eIF4E. Esta interacción aproxima el extremo 5′ y el extremo 3′ formando un lazo cerrado que estabiliza la unión ribosomal y facilita ciclos sucesivos de traducción.
12.3.3. Poliadenilación alternativa
Cuando un gen tiene múltiples señales AAUAAA, la selección de uno u otro sitio de corte determina la longitud del extremo 3′ no traducido (3′ UTR) y, en algunos casos, si se incluyen o excluyen exones codificantes.
Un 3′ UTR largo contiene más sitios de unión para miARN y proteínas reguladoras, lo que tiende a reducir la estabilidad y la traducción del ARNm. La poliadenilación alternativa es, por tanto, un mecanismo de regulación de la expresión génica independiente del splicing.
12.4. Splicing: eliminación de intrones
12.4.1. Estructura del intrón: secuencias consenso
Los genes eucariotas están interrumpidos por intrones intercalados entre exones. El splicing elimina los intrones y une los exones para formar el ARNm maduro. Para que el spliceosoma reconozca un intrón correctamente, este debe presentar tres elementos de secuencia conservados:
- El sitio donador o 5′ ss es la secuencia GU en el extremo 5′ del intrón.
- El sitio aceptor o 3′ ss es la secuencia AG en el extremo 3′, precedido por un tracto de pirimidinas.
- El punto de rama es una adenosina conservada situada entre 18 y 40 nucleótidos antes del sitio aceptor, que participa directamente en la primera reacción química.
Regla GU-AG (regla de Chambon): todos los intrones tipo U2 comienzan con GU y terminan con AG. Las mutaciones en estas posiciones invariantes causan splicing aberrante y son causa frecuente de enfermedades monogénicas.
12.4.2. Mecanismo molecular: el spliceosoma y las dos transesterificaciones
El splicing es catalizado por el spliceosoma, un complejo ribonucleoproteico dinámico formado por cinco snRNP: U1, U2, U4, U5 y U6. Cada snRNP contiene un ARN nuclear pequeño (snARN) asociado a proteínas; el snARN aporta la capacidad de reconocimiento por complementariedad con las secuencias consenso del intrón.
El spliceosoma se ensambla de forma ordenada sobre el pre-ARNm en varios pasos.
- U1 reconoce el sitio 5′ de splicing.
- U2 reconoce el punto de rama.
- El tri-snRNP U4/U6·U5 se incorpora y reposiciona los sitios reactivos.
La reacción química del splicing ocurre en dos transesterificaciones sucesivas.
- Primera transesterificación: el grupo 2′-OH de la adenosina del punto de rama ataca el enlace fosfodiéster del sitio 5′ ss. Se forma una estructura en lazo denominada lariat: el extremo 5′ del intrón queda unido covalentemente a la adenosina del punto de rama mediante un enlace 2′–5′, y el exón 5′ queda con un extremo 3′-OH libre.
- Segunda transesterificación: el extremo 3′-OH libre del exón 5′ ataca el enlace fosfodiéster del sitio 3′ ss. Los dos exones quedan unidos y el intrón se libera en forma de lariat, que es degradado en el núcleo.
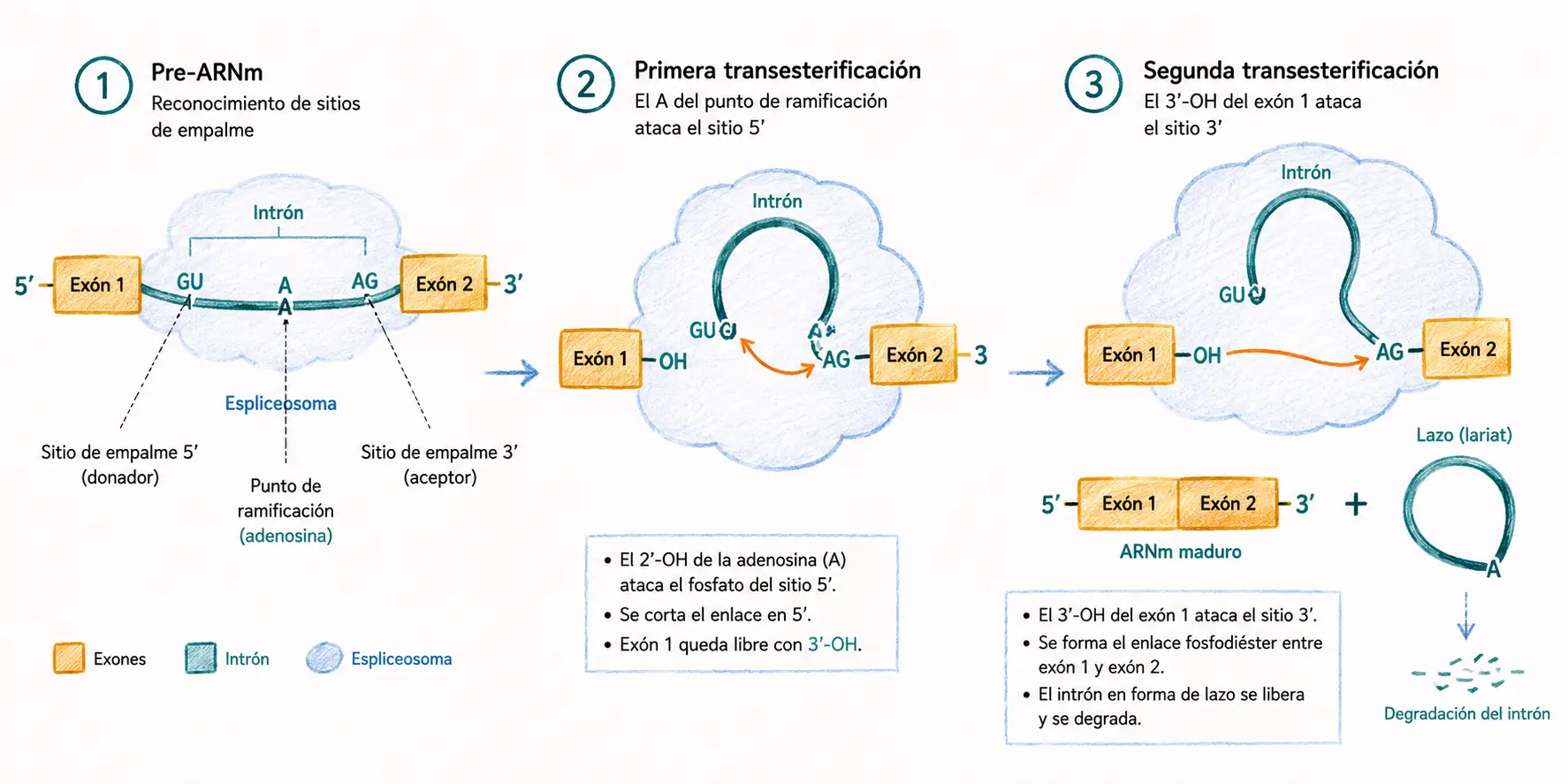
El splicing no consume energía neta: el número de enlaces fosfodiéster se conserva. Las transesterificaciones son reacciones de intercambio, no de hidrólisis.
12.4.3. Splicing alternativo: concepto y patrones principales
En la mayoría de los genes humanos multi-exónicos, la selección de qué secuencias se incluyen como exones puede variar según el tipo celular, el estadio de desarrollo o las condiciones fisiológicas. Se calcula que más del 90% de los genes humanos con más de un exón sufren splicing alternativo, lo que permite generar entre 80.000 y 100.000 proteínas distintas a partir de unos 20.000 genes.
Los patrones principales son cuatro:
- Exon skipping: es el más frecuente en mamíferos: un exón interno se omite y los exones flanqueantes se unen directamente.
- Uso de sitios alternativos 5′ o 3′: genera variantes con secuencias ligeramente distintas en los extremos de los exones.
- Retención de intrón: ocurre cuando un intrón no se elimina y queda incluido en el ARNm maduro; es poco frecuente en mamíferos pero habitual en plantas.
- Inclusión de exón alternativo: incorpora un exón que en otras condiciones se omite.
Las mutaciones en los sitios consenso de splicing (posiciones GU y AG, y el punto de rama) causan splicing aberrante y son responsables de enfermedades monogénicas bien caracterizadas: beta-talasemia, algunas formas de fibrosis quística (mutación 3849+10kb C>T en CFTR) y atrofia muscular espinal (por splicing aberrante del gen SMN2).
12.5. Edición del ARN
12.5.1. Mecanismos principales: edición C→U (APOBEC) y A→I (ADAR)
La edición del ARN es el proceso por el cual se alteran nucleótidos individuales en la secuencia de un ARN transcrito sin modificar el ADN. Es postranscripcional y no heredable.
Los dos mecanismos principales en mamíferos son deaminaciones:
- La edición C→U está catalizada por la familia APOBEC. La deaminación convierte citosina en uracilo, que el ribosoma lee como timina. El ejemplo canónico es el ARNm de la apolipoproteína B (ApoB): en el hígado se produce la isoforma larga ApoB-100 sin edición; en el intestino, APOBEC1 edita el codón CAA (glutamina) en UAA (codón de parada), generando la isoforma truncada ApoB-48, necesaria para el ensamblaje de quilomicrones.
- La edición A→I está catalizada por las enzimas ADAR. La deaminación convierte adenosina en inosina, que el ribosoma interpreta como guanosina. El ejemplo más estudiado es el receptor de glutamato tipo AMPA en el sistema nervioso central: la edición A→I en la subunidad GluA2 cambia el aminoácido en la región del canal iónico, reduciendo drásticamente la permeabilidad al calcio. En condiciones normales, prácticamente el 100% de los ARNm de GluA2 están editados en esta posición en el cerebro adulto.
Edición de GluA2 y excitotoxicidad: la pérdida de edición A→I en GluA2 convierte el canal AMPA en permeable al calcio, favoreciendo la excitotoxicidad neuronal. Su alteración se ha asociado a glioblastoma y algunas formas de esclerosis lateral amiotrófica. Por otro lado, la actividad APOBEC desregulada sobre el ADN genómico propio es uno de los patrones de mutagénesis más frecuentes en carcinomas de mama y pulmón.
12.6. Estabilidad y degradación del ARNm
12.6.1. Determinantes de la vida media del ARNm
Los ARNm no son moléculas permanentes. Su vida media varía desde minutos (ARNm de citocinas o proto-oncogenes) hasta horas o días (ARNm de proteínas estructurales como la beta-globina). Los principales determinantes son tres:
- Estado del cap 5′: mientras el cap está intacto, la exonucleasa Xrn1 no puede atacar el extremo 5′.
- Longitud de la cola poli(A): a medida que la cola se acorta por deadenilasas, el ARNm pierde su escudo 3′ y su capacidad de circularizarse.
- Elementos AU-rich (ARE): secuencias AUUUA repetidas en el 3′ UTR que reclutan proteínas desestabilizadoras. Los ARNm de citocinas proinflamatorias (TNF-α, IL-6) y de proto-oncogenes (c-fos, c-myc) están cargados de AREs, garantizando que su expresión sea rápidamente reversible.
12.6.2. Vías de degradación del ARNm
La vía mayoritaria de degradación del ARNm en eucariotas comienza con:
- Deadenilación por el complejo CCR4-NOT, que acorta progresivamente la cola poli(A)..
- Decapping: el complejo DCP1/DCP2 elimina el cap m7G del extremo 5′ cuando la cola cae por debajo de ~10–15 adenosinas.
- Degradación por la exonucleasa Xrn1: el ARNm descapado queda expuesto a la Xrn1 que lo degrada de forma procesiva en dirección 5’→3′.
De forma alternativa, el extremo 3′ descubierto puede ser atacado por el exosoma, complejo multiproteico con actividad exonucleásica 3′→5′.
12.6.3. Vigilancia de calidad del ARNm: el NMD
El NMD (nonsense-mediated mRNA decay) detecta y elimina ARNm con codones de parada prematuros (PTC). Un PTC puede surgir de mutaciones sin sentido, errores de splicing o cambios de marco de lectura.
El NMD aprovecha el complejo de unión a la unión exón-exón (EJC), depositado por el spliceosoma a unos 20–24 nucleótidos aguas arriba de cada sitio de unión exón-exón. En condiciones normales, el ribosoma desplaza los EJC durante la primera ronda de traducción. Si el ribosoma encuentra un codón de parada y todavía queda al menos un EJC aguas abajo, el complejo UPF1/UPF2/UPF3 desencadena la degradación acelerada del ARNm.
NMD y enfermedades monogénicas: aproximadamente el 30% de las mutaciones causantes de enfermedades monogénicas generan codones de parada prematuros. El NMD elimina ese ARNm, resultando en pérdida total de función. Fármacos como el atalurén pueden suprimir la lectura del PTC y restaurar parcialmente la producción de proteína funcional.
12.7. Regulación por ARN no codificantes pequeños
12.7.1. miARN: biogénesis y mecanismo de acción
Los microARN (miARN) son ARN no codificantes de aproximadamente 22 nucleótidos que reprimen la expresión génica postranscripcionalmente. Se codifican en el genoma y se procesan en dos etapas: una nuclear y una citoplásmica.
En el núcleo, la ARN polimerasa II transcribe el gen del miARN generando un transcrito largo denominado pri-miARN, que forma una estructura de horquilla. El complejo Drosha/DGCR8 reconoce y corta esa horquilla liberando un precursor de ~60–70 nucleótidos con estructura de tallo-bucle: el pre-miARN. El pre-miARN es exportado al citoplasma por Exportina-5.
En el citoplasma, la endonucleasa Dicer corta el tallo del pre-miARN generando un dúplex de ~22 pb. Una de las dos cadenas (la cadena guía) se carga en la proteína Argonauta (AGO2) formando el complejo RISC (RNA-induced silencing complex). La cadena pasajera se degrada.
El RISC cargado con el miARN busca ARNm con secuencias complementarias en el 3′ UTR. La complementariedad suele ser parcial, especialmente en la región semilla (nucleótidos 2–8 desde el extremo 5′). Si la complementariedad es casi perfecta, AGO2 escinde el ARNm; si es parcial, el RISC bloquea la traducción y promueve la deadenilación y degradación del ARNm.
Un mismo miARN puede regular cientos de ARNm distintos, y un mismo ARNm puede ser regulado por múltiples miARN. Esta promiscuidad no es un defecto: permite coordinar redes reguladoras complejas de forma coordinada.
12.7.2. ARNsi: mecanismo y diferencia con miARN
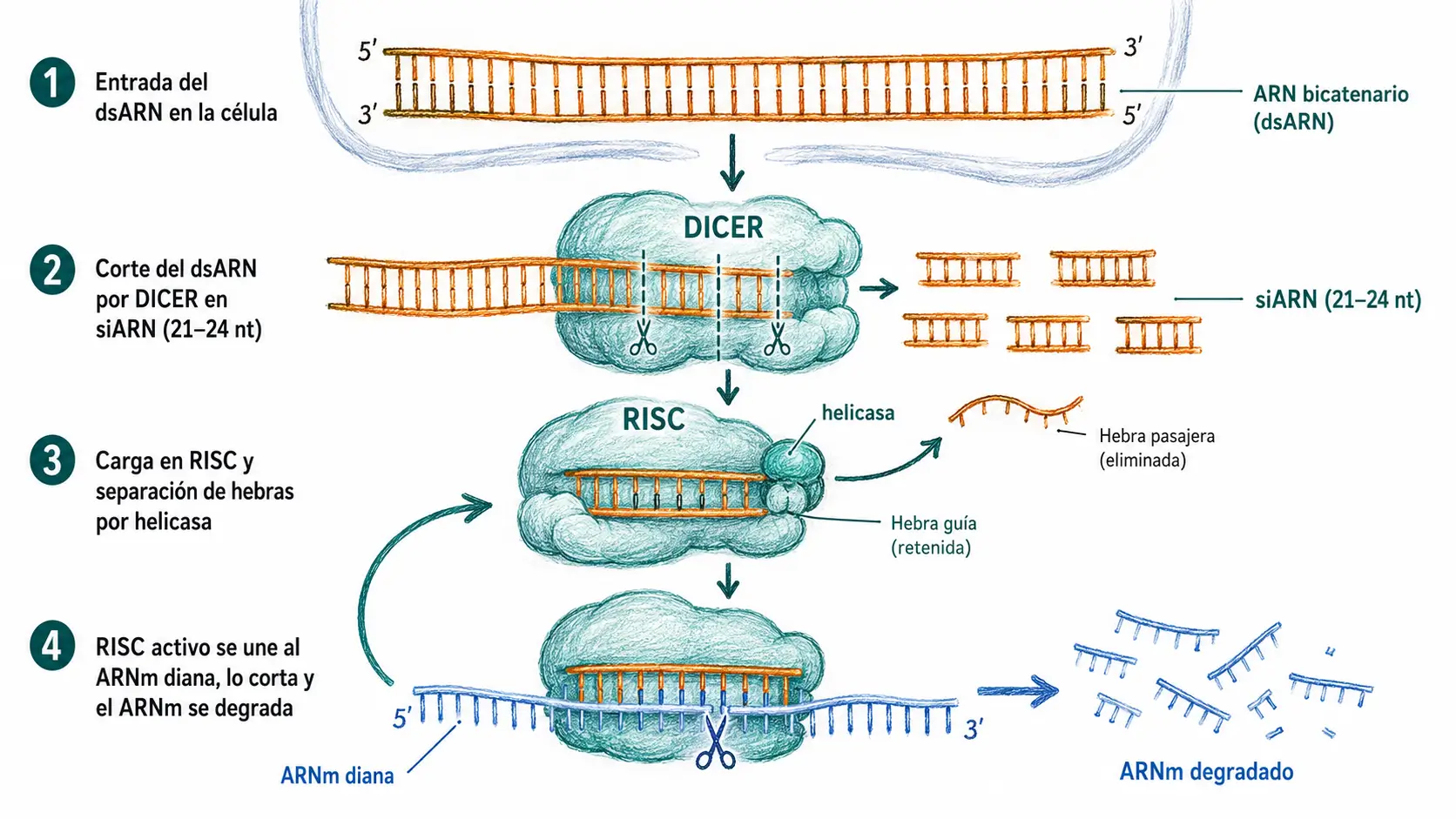
Los ARN de interferencia pequeños (ARNsi) comparten la maquinaria efectora (RISC con Argonauta) con los miARN, pero difieren en origen, procesamiento y complementariedad con la diana.
Los ARNsi proceden de ARN de doble cadena exógeno, habitualmente viral, o de dsARN sintético introducido experimentalmente. Dicer los procesa directamente sin necesidad del paso nuclear de Drosha. La diferencia funcional más importante es que los ARNsi tienen complementariedad perfecta con su diana: esto provoca siempre el corte endonucleolítico del ARNm por AGO2. No bloquean la traducción: destruyen el ARNm.
| Característica | miARN | ARNsi |
|---|---|---|
| Origen | Genoma propio (gen de miARN) | dsARN exógeno (viral) o sintético |
| Procesamiento nuclear | Sí: Drosha/DGCR8 genera el pre-miARN | No: Dicer procesa directamente el dsARN |
| Complementariedad con la diana | Parcial (región semilla, nt 2–8) | Perfecta |
| Mecanismo de silenciamiento | Bloqueo de traducción + deadenilación | Corte endonucleolítico del ARNm por AGO2 |
| Número de dianas | Múltiples (cientos de ARNm) | Una diana específica por ARNsi |
| Complejo efector | RISC (con Argonauta) | RISC (con Argonauta) |
| Función biológica principal | Regulación génica endógena | Defensa antiviral; herramienta terapéutica |
Patisirán y ARN de interferencia terapéutico: patisirán fue el primer ARNsi terapéutico aprobado. Resolvió el problema histórico de la entrega intracelular mediante nanopartículas lipídicas ionizables, la misma plataforma utilizada posteriormente para las vacunas de ARNm frente a SARS-CoV-2.
Los lncARN (ARN no codificantes largos, >200 nucleótidos) regulan la expresión génica a múltiples niveles, incluyendo la estabilidad del ARNm y la traducción. Su biología está siendo activamente investigada y su desarrollo detallado excede el alcance de este tema.
