10.1. Características generales de la replicación del ADN
La replicación del ADN es semiconservativa: tras completarse, cada doble hélice hija contiene una hebra original parental y una de nueva síntesis. El proceso es bidireccional, ya que la síntesis avanza en dirección 5′ a 3′ en cada una de las dos hebras antiparalelas, y completo, porque se replica la totalidad del ADN. La replicación está además ligada al ciclo celular: en condiciones normales el ADN solo se replica cuando la célula se va a dividir, aunque existen excepciones a esta regla.
Replicón: la unidad de ADN que se replica a partir de un único origen de replicación. En procariotas existe un único origen de replicación (el OriC en E. coli), mientras que en eucariotas hay múltiples orígenes, entre 10³ y 10⁴.
La replicación requiere mecanismos que garanticen simultáneamente velocidad y fidelidad, para no cometer errores en la copia. Interviene un gran número de proteínas, siendo la ADN polimerasa III la responsable principal de la síntesis del ADN en procariotas, y conlleva un elevado gasto energético.
| Característica | Significado | Implicación funcional |
|---|---|---|
| Semiconservativa | Cada molécula hija conserva una hebra parental y sintetiza una nueva | Garantiza la transmisión fiel de la información genética a las células hijas |
| Bidireccional | Dos horquillas de replicación avanzan en sentidos opuestos desde el origen | Reduce el tiempo necesario para replicar el cromosoma completo |
| Semidiscontinua | La hebra conductora se sintetiza de forma continua; la retardada, en fragmentos de Okazaki | Consecuencia directa de que todas las polimerasas sintetizan en sentido 5'→3' |
| Completa | Se replica la totalidad del ADN antes de la división celular | La regulación se ejerce sobre el inicio, no sobre la elongación |
| Ligada al ciclo celular | En condiciones normales, el ADN solo se replica cuando la célula se va a dividir | Evita la replicación descontrolada; su pérdida es un rasgo frecuente en células cancerosas |
| Compleja y de alta fidelidad | Intervienen múltiples proteínas con mecanismos de corrección de errores | Tasa de error de 10⁻⁸ a 10⁻⁹ gracias a la actividad exonucleásica 3'→5' |
10.2. Características de las ADN polimerasas
Las ADN polimerasas sintetizan ADN de forma dirigida por una hebra molde, requieren un cebador de ARN con extremo 3′-OH libre para comenzar, y solo pueden elongar en dirección 5′ a 3′. La reacción añade un desoxinucleótido trifosfato (dNTP) a la cadena naciente con liberación de pirofosfato: (DNA)ⁿ + dNTP → (DNA)ⁿ⁺¹ + PPi. La hidrólisis posterior del PPi por las pirofosfatasas hace la reacción termodinámicamente irreversible.
Además de la actividad polimerizadora, las ADN polimerasas poseen actividad exonucleásica en dirección 3′ a 5′, que permite la autocorrección: si se incorpora un nucleótido incorrecto, la enzima lo extrae antes de continuar la elongación. La ADN polimerasa I posee adicionalmente actividad exonucleásica en dirección 5′ a 3′, lo que le permite eliminar los cebadores de ARN durante la replicación y la reparación.
En E. coli existen tres ADN polimerasas principales. La ADN polimerasa I participa en la reparación y en la replicación. La ADN polimerasa II tiene una función poco conocida, probablemente relacionada con la reparación del ADN. La ADN polimerasa III es la responsable de la síntesis del ADN en la replicación y actúa como holoenzima.
| Polimerasa | Síntesis 5'→3' | Exonucleasa 3'→5' | Exonucleasa 5'→3' | Función principal |
|---|---|---|---|---|
| Pol I | Sí | Sí | Sí | Eliminación de cebadores de ARN; reparación y síntesis de relleno |
| Pol II | Sí | Sí | No | Reparación del ADN (función poco caracterizada) |
| Pol III | Sí | Sí | No | Síntesis principal del ADN en la replicación; alta procesividad |
Una holoenzima es la forma completa y activa de una enzima, compuesta por la apoenzima y su cofactor, necesaria para catalizar reacciones bioquímicas.
La procesividad de la ADN polimerasa III, es decir, su capacidad de no disociarse del molde durante la elongación, le permite alcanzar una velocidad de síntesis de 1000 nucleótidos por segundo en procariotas (frente a 100 nucleótidos por segundo en eucariotas). La elongación requiere molde, cebador, dNTP y Mg²+. La tasa de error es de 10⁻⁸ a 10⁻⁹ gracias a la actividad exonucleásica 3' a 5' de autocorrección.
10.3. Proteínas de la replicación y mecanismo de inicio en procariotas: el modelo E. coli
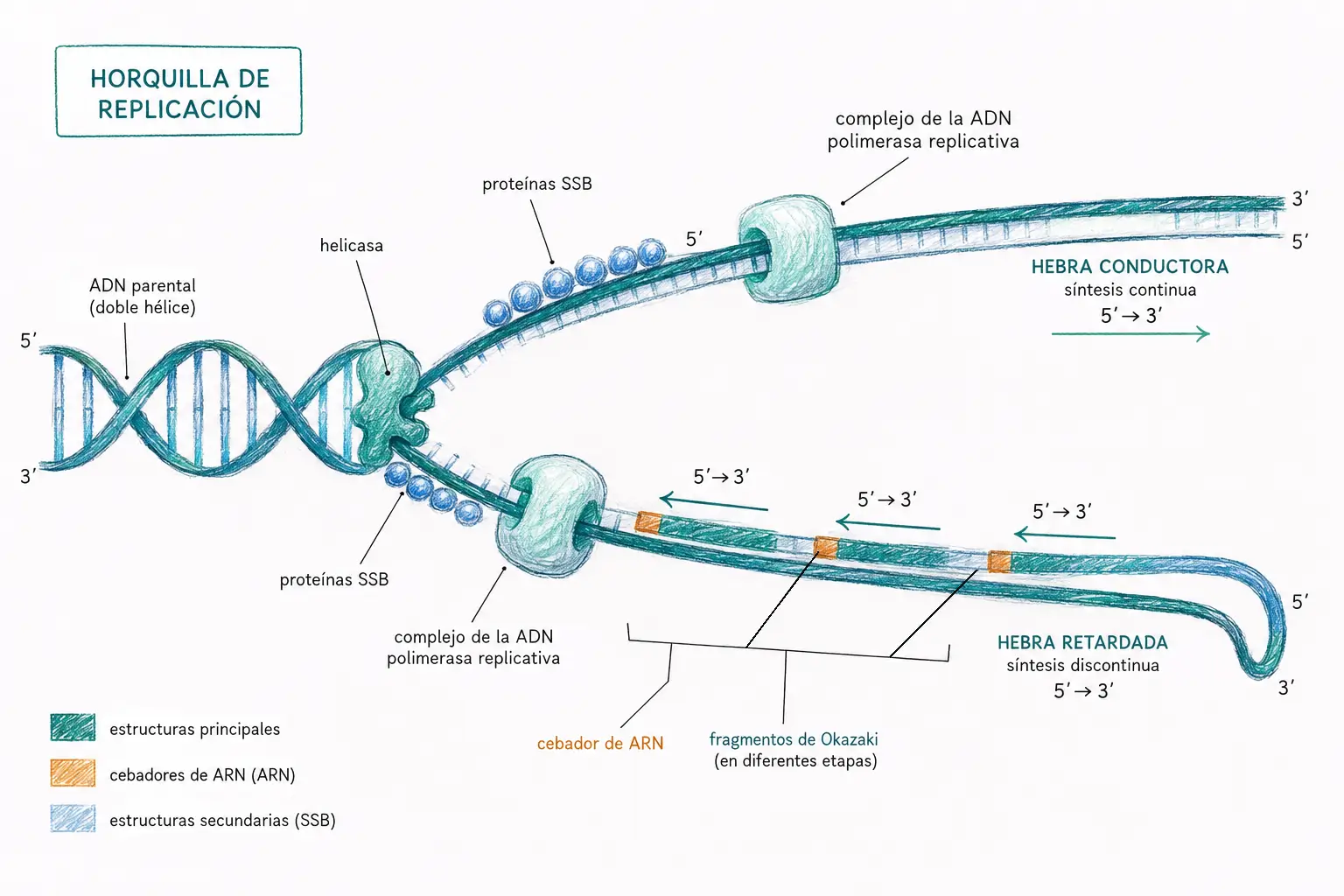
La replicación en E. coli requiere, además de las ADN polimerasas, un conjunto coordinado de proteínas especializadas.
- Las proteínas DnaA, que actúan como tetrámeros, localizan y se unen al OriC.
- Las helicasas (DnaB, DnaC y la proteína Rep) abren la doble hélice rompiendo los puentes de hidrógeno entre las hebras.
- Las proteínas SSB estabilizan el ADN monocatenario recién separado.
- La primasa DnaG, una ARN polimerasa ADN-dependiente, sintetiza los cebadores de ARN necesarios para que la ADN polimerasa III pueda comenzar la síntesis.
- Las ADN ligasas sellan las interrupciones en la cadena nueva.
- La ADN girasa (topoisomerasa II) elimina los súperenrollamientos positivos que se acumulan por delante de la horquilla.
- Las proteínas Tus se unen a los genes de terminación ter y detienen el avance de la horquilla, de modo que la replicación termina por acción de secuencias génicas específicas y no por el cruce físico de las dos horquillas.
Para que la replicación pueda iniciarse en el OriC, el ADN debe estar no metilado y súperenrollado negativamente. Las proteínas DnaA reconocen secuencias específicas del OriC y se unen a ellas de forma cooperativa: cada unión facilita la siguiente, formando un agregado que tensa la región OriC y provoca la apertura de la doble hélice. La apertura ocurre en una zona rica en secuencias A=T repetitivas, que favorecen la separación de hebras al tener solo dos puentes de hidrógeno. Al ser la replicación completa, la regulación se ejerce exclusivamente a nivel del inicio.
Una vez abierta la doble hélice, las helicasas amplían progresivamente la zona de separación mientras las DnaA se disocian del ADN. Las proteínas SSB estabilizan el ADN monocatenario, las girasas eliminan los súperenrollamientos positivos, la primasa fabrica los cebadores y la ADN polimerasa III comienza la síntesis. En la hebra retardada, la maquinaria de replicación avanza sobre el ADN sin copiarlo hasta que la primasa sintetiza un nuevo cebador de ARN cada 1000 a 2000 nucleótidos, que se estabiliza con proteínas SSB. El ADN de la hebra retardada se invierte en un bucle para que ambas hebras se sinteticen de forma coordinada en la misma horquilla.
10.4. Replicación en eucariotas
La replicación eucariota guarda grandes similitudes con la procariota, pero es considerablemente más compleja y está menos caracterizada en detalle.
La diferencia más relevante es la presencia de múltiples orígenes de replicación, entre 10³ y 10⁴, que no se activan todos simultáneamente: existe una secuencia de activación establecida en la que los orígenes situados en eucromatina, la cromatina activa, se activan antes durante la fase S, y los demás se incorporan progresivamente. La velocidad de síntesis es menor que en procariotas, 50 nucleótidos por segundo, y los fragmentos de Okazaki son más cortos, produciéndose uno cada 100 nucleótidos.
En eucariotas existen cinco ADN polimerasas principales con funciones diferenciadas.
- La ADN polimerasa α sintetiza la hebra retardada.
- La ADN polimerasa β participa en la reparación.
- La ADN polimerasa γ replica el ADN mitocondrial.
- La ADN polimerasa δ sintetiza la hebra continua formando complejo con el PCNA (antígeno nuclear de las células en proliferación), equivalente funcional de la subunidad β de la ADN polimerasa III de E. coli.
- La ADN polimerasa ε participa tanto en la replicación como en la reparación.
| Polimerasa | Función principal | Observaciones |
|---|---|---|
| Pol α | Síntesis de la hebra retardada | Asociada a actividad primasa; inicia la síntesis de los fragmentos de Okazaki |
| Pol β | Reparación del ADN | No interviene en la replicación ordinaria |
| Pol γ | Replicación del ADN mitocondrial | Única polimerasa presente en la mitocondria |
| Pol δ | Síntesis de la hebra continua | Forma complejo con PCNA, equivalente funcional de la subunidad β de Pol III |
| Pol ε | Replicación y reparación | Función dual; participa en la síntesis de la hebra adelantada en algunos modelos |
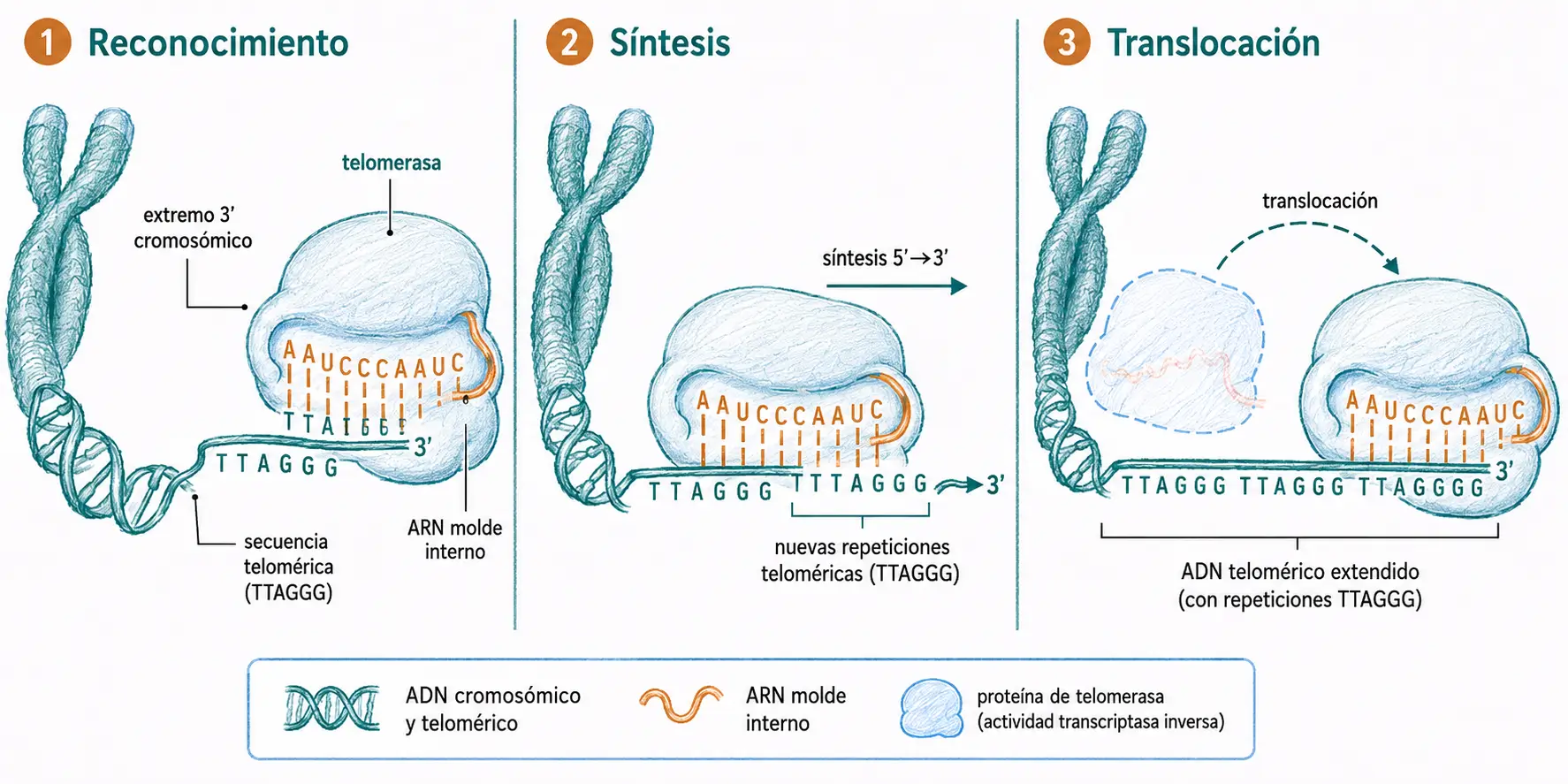
Las telomerasas resuelven el problema de la replicación de los extremos cromosómicos. Son riboproteínas formadas por una parte proteica y una molécula de ARN cuya secuencia es copia de la secuencia repetida del telómero. Actuando como transcriptasas inversas, copian su propio ARN molde en ADN y lo añaden al extremo 3′ del cromosoma, generando así un nuevo punto de inicio a partir del cual se completa la replicación del extremo
En las células cancerosas la expresión de la telomerasa está aumentada, lo que permite eludir el acortamiento telomérico y proliferar de forma indefinida. Por este motivo, la inhibición de la telomerasa es una diana terapéutica activa en oncología.
La unión del ADN a las histonas para formar nucleosomas añade una capa adicional de complejidad a la replicación eucariota: la cromatina debe desensamblarse por delante de la horquilla y volver a ensamblarse tras el paso de la maquinaria replicativa. El control del inicio de la replicación en eucariotas sigue los mismos principios generales que en procariotas, ejerciéndose sobre el inicio y no sobre la elongación.
10.5. Reacción en cadena de la polimerasa (PCR)
La PCR es una técnica de amplificación enzimática in vitro de fragmentos específicos de ADN mediante ciclos repetidos de desnaturalización, hibridación de cebadores y extensión. Cada ciclo duplica teóricamente la cantidad de ADN molde, lo que permite generar millones de copias de un fragmento específico en pocas horas a partir de cantidades mínimas de material de partida.
El protocolo básico consiste en tres pasos por ciclo:
- Calentar la solución de ADN con una ADN polimerasa termorresistente a 95°C durante 15 segundos para desnaturalizar la doble hélice.
- Bajar la temperatura a 55°C durante 1 minuto para que los cebadores específicos se hibriden con sus secuencias complementarias en cada hebra.
- Elevar la temperatura a 72°C durante 30 segundos para que la polimerasa extienda los cebadores y sintetice las cadenas nuevas.
Las aplicaciones clínicas de la PCR son prácticamente ilimitadas: diagnóstico de enfermedades infecciosas, detección de mutaciones, genotipado, medicina forense y diagnóstico prenatal, entre otras. El principal problema técnico es la contaminación de la muestra, que puede generar resultados falsos positivos.
