14.1. Introducción: del polipéptido a la proteína funcional
La cadena polipeptídica que emerge del ribosoma es una secuencia lineal de aminoácidos. En la gran mayoría de los casos, esta cadena no es funcional en ese estado: debe plegarse en su conformación tridimensional activa, dirigirse al compartimento subcelular correcto, y sufrir una serie de modificaciones químicas que ajustan su actividad, estabilidad e interacciones.
El conjunto de estas transformaciones se denomina modificaciones post-traduccionales (MPT). Son tan diversas y específicas que el número de proteínas funcionalmente distintas que puede generar el genoma humano supera en varios órdenes de magnitud el número de genes codificantes.
Las modificaciones post-traduccionales no son pasos pasivos: son mecanismos de regulación activa. La adición o eliminación de un grupo fosfato, un grupo acetilo o un azúcar puede activar o inactivar una proteína en segundos, sin necesidad de nueva transcripción ni traducción. Esto las convierte en el nivel de respuesta más rápido de la regulación de la expresión génica.
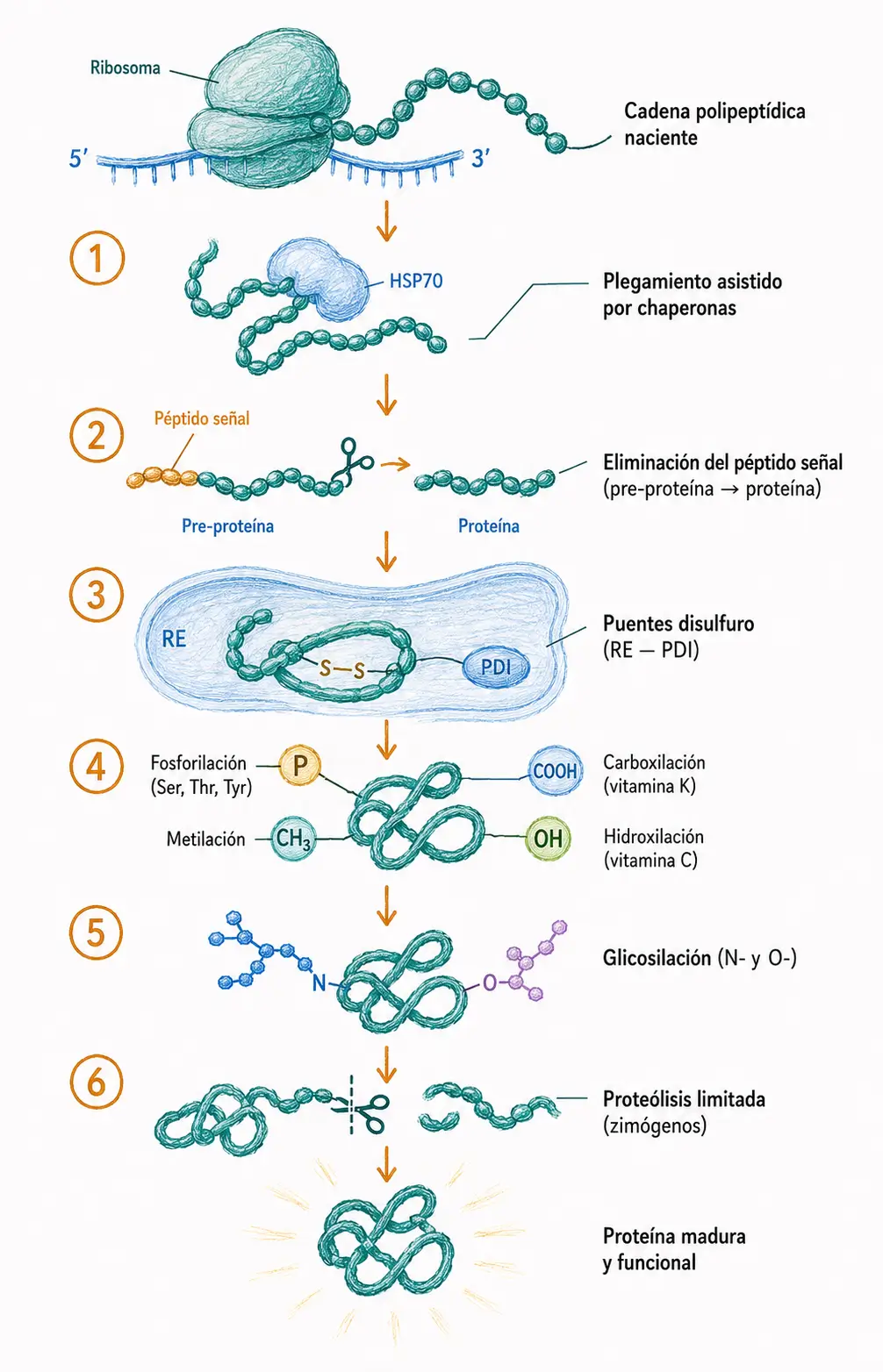
14.2. Plegamiento y chaperonas
La información necesaria para que una proteína adopte su conformación tridimensional nativa está contenida en su secuencia de aminoácidos: este es el principio de Anfinsen, establecido experimentalmente con la ribonucleasa A. Sin embargo, la célula es un medio muy concentrado en proteínas, y las cadenas polipeptídicas nacientes exponen regiones hidrofóbicas que tienden a agregarse con otras cadenas antes de completar su plegamiento.
Para evitar estas agregaciones, las células disponen de chaperonas moleculares: proteínas que reconocen y se unen transitoriamente a regiones hidrofóbicas expuestas, previniendo interacciones incorrectas mientras la cadena termina de sintetizarse o se recupera de un estrés. Las chaperonas no aportan información sobre el plegamiento correcto, solo crean las condiciones para que ocurra.
Las principales familias de chaperonas en eucariotas son las HSP70 (heat shock proteins de 70 kDa), que acompañan al polipéptido naciente desde el ribosoma, y las chaperonoínas HSP60 (complejo GroEL/GroES en procariotas), que forman cámaras proteicas donde el polipéptido puede plegarse aislado del resto del citoplasma.
Las chaperonas se sobreexpresan en respuesta a estrés térmico, oxidativo o hipóxico, situaciones en las que las proteínas tienden a desplegarse. En patología, la acumulación de proteínas mal plegadas que escapan al control de las chaperonas y forman agregados insolubles es el mecanismo central de las enfermedades por depósito de proteínas mal plegadas (misfolding diseases), como la enfermedad de Alzheimer (agregados de tau y β-amiloide), el Parkinson (α-sinucleína) o las encefalopatías espongiformes priónicas.
14.3. Modificaciones de los extremos de la cadena
14.3.1. Eliminación de la metionina o formilmetionina inicial
Toda cadena polipeptídica comienza con metionina (eucariotas) o formilmetionina (procariotas) como consecuencia del codón de inicio AUG. Sin embargo, la mayoría de las proteínas maduras no conservan este residuo N-terminal: una metionina aminopeptidasa lo elimina cotranslacionalmente en aproximadamente el 50–70% de las proteínas, dependiendo del residuo que le sigue.
14.3.2. Acetilación del extremo N-terminal
Tras la eliminación de la metionina inicial, el nuevo extremo N-terminal puede ser acetilado por N-acetiltransferasas citoplásmicas. La acetilación N-terminal es una de las modificaciones más frecuentes en proteínas eucariotas (afecta a más del 80% del proteoma humano), y contribuye a la estabilidad proteica y a interacciones proteína-proteína. También regula la vida media de las proteínas según la regla del extremo N-terminal (regla N-end): ciertos residuos N-terminales actúan como señales de degradación por el proteasoma.
14.4. Puentes disulfuro
Los puentes disulfuro son enlaces covalentes que se forman entre los grupos tiol (-SH) de dos residuos de cisteína. Estabilizan enormemente la estructura terciaria y cuaternaria de las proteínas, especialmente las que se secretan al exterior o se anclan en la membrana plasmática, que deben resistir el ambiente extracelular sin el apoyo de chaperonas.
La formación de puentes disulfuro requiere un ambiente oxidante, por lo que ocurre en el retículo endoplasmático (RE), donde la enzima proteína disulfuro isomerasa (PDI) cataliza tanto la formación como el reordenamiento de puentes disulfuro incorrectos hasta alcanzar la configuración nativa. El citoplasma, por ser reductor, no permite la formación de puentes disulfuro estables.
Los agentes reductores como el β-mercaptoetanol o el DTT rompen los puentes disulfuro, desnaturalizando las proteínas que dependen de ellos para su estructura. Esto tiene aplicación directa en electroforesis en condiciones desnaturalizantes (SDS-PAGE reductora) y en técnicas de bioquímica proteica.
14.5. Modificaciones covalentes de residuos laterales
14.5.1. Fosforilación
La fosforilación es la modificación post-traduccional reversible más importante en la regulación de la actividad proteica. Consiste en la transferencia de un grupo fosfato del ATP a un residuo hidroxilado de la proteína: serina, treonina o tirosina.
La cataliza una familia de enzimas denominadas quinasas o kinasas (adición del fosfato) y se revierte por fosfatasas (eliminación del fosfato). El cambio de carga que introduce el grupo fosfato (−2 a pH fisiológico) altera las interacciones electrostáticas de la proteína y produce cambios conformacionales que modifican su actividad, sus interacciones con otras proteínas o su localización subcelular.
La desregulación de la fosforilación es uno de los mecanismos centrales del cáncer. Muchos oncogenes codifican tirosina quinasas constitutivamente activas (Bcr-Abl en la leucemia mieloide crónica, EGFR en cáncer de pulmón, HER2 en cáncer de mama) que mantienen señales proliferativas encendidas de forma permanente. Los inhibidores de tirosina quinasas (imatinib, erlotinib, trastuzumab) son los fármacos diana más exitosos de la oncología molecular.
14.5.2. Metilación
La metilación de residuos de lisina o arginina es frecuente en proteínas musculares como la actina y la miosina. Los grupos metilo introducen zonas hidrofóbicas que favorecen interacciones específicas entre proteínas. En el contexto de las histonas, la metilación de lisinas específicas es una marca epigenética que puede activar o reprimir la transcripción dependiendo de la posición y el grado de metilación.
14.5.3. Carboxilación
La carboxilación de residuos de glutamato añade un grupo carboxilo adicional en la posición γ, generando γ-carboxiglutamato (Gla). Esta modificación depende de la vitamina K como cofactor y es esencial para los factores de coagulación vitamina K-dependientes (protrombina (factor II), factor VII, factor IX y factor X).
Los residuos Gla tienen alta afinidad por el Ca²⁺: su quelación posiciona los factores de coagulación sobre las superficies de membranas fosfolipídicas cargadas negativamente, donde ocurre la cascada de coagulación.
Los anticoagulantes cumarínicos (warfarina, acenocumarol) actúan inhibiendo la vitamina K epóxido reductasa, bloqueando el ciclo de la vitamina K e impidiendo la carboxilación de los factores de coagulación. El resultado es la producción de factores sin residuos Gla, incapaces de unir Ca²⁺ y funcionalmente inactivos. Este es el mecanismo de acción de los anticoagulantes orales clásicos.
14.5.4. Hidroxilación
La hidroxilación de residuos de prolina y lisina en el colágeno, catalizada por las enzimas prolil 4-hidroxilasa y lisil hidroxilasa con vitamina C como cofactor esencial, introduce grupos -OH que permiten la formación de puentes de hidrógeno entre las tres cadenas de la triple hélice del colágeno, estabilizando enormemente la estructura y proporcionando resistencia mecánica al tejido conjuntivo.
El escorbuto es la consecuencia de la deficiencia de vitamina C: sin vitamina C, la prolil hidroxilasa no puede catalizar la reacción, el colágeno sintetizado carece de hidroxiprolina y es mecánicamente inestable. Las manifestaciones clínicas (fragilidad vascular, hemorragias, mala cicatrización) reflejan directamente el papel estructural del colágeno en el tejido conjuntivo.
14.6. Glicosilación y lipidación
14.6.1. Glicosilación
La glicosilación es la adición covalente de cadenas de azúcares (oligosacáridos) a la proteína. Es la modificación post-traduccional más compleja en términos de diversidad estructural.
Existen dos tipos principales:
- La N-glicosilación ocurre en el retículo endoplasmático: el oligosacárido se transfiere en bloque a la cadena lateral de una asparragina (Asn) en la secuencia consenso Asn-X-Ser/Thr.
- La O-glicosilación ocurre en el aparato de Golgi: el azúcar se añade al grupo -OH de serina o treonina, sin secuencia consenso estricta.
Las glicoproteínas son ubicuas en la superficie celular y en la matriz extracelular. Sus funciones incluyen el reconocimiento celular (grupos sanguíneos ABO), la respuesta inmune (anticuerpos IgG son glicoproteínas), la protección mecánica de mucosas (mucinas) y el aumento de la estabilidad y solubilidad de proteínas secretadas.
14.6.2. Lipidación
La lipidación es la adición covalente de grupos lipídicos a proteínas, lo que les confiere afinidad por membranas. Los principales tipos son:
- La miristoilación (ácido mirístico en Gly N-terminal).
- La palmitoilación (ácido palmítico en Cys).
- La prenilación (grupos farnesilo o geranilgeranilo en Cys C-terminal).
Muchas proteínas de señalización intracelular (proteínas G, Ras) requieren lipidación para anclarse a la cara interna de la membrana plasmática y ejercer su función.
| Modificación | Residuo diana | Cofactor / enzima | Función / ejemplo clínico |
|---|---|---|---|
| Fosforilación | Ser, Thr, Tyr | Quinasas / Fosfatasas (reversible) | Regulación de señalización; quinasas oncogénicas (Bcr-Abl, EGFR) |
| Acetilación N-terminal | Gly / Met N-terminal | N-acetiltransferasas | Estabilidad proteica; regla N-end (degradación proteasomal) |
| Metilación | Lys, Arg | Metiltransferasas (SAM como donador) | Interacciones hidrofóbicas (actina, miosina); epigenética (histonas) |
| Carboxilación (γ-Gla) | Glu | γ-carboxilasa (vitamina K) | Unión de Ca²⁺; factores de coagulación (protrombina, VII, IX, X); warfarina |
| Hidroxilación | Pro, Lys | Prolil/lisil hidroxilasa (vitamina C) | Estabilidad del colágeno; escorbuto por déficit de vitamina C |
| Puentes disulfuro | Cys – Cys | PDI (retículo endoplasmático) | Estabilidad estructural de proteínas secretadas y de membrana |
| N-glicosilación | Asn (secuencia Asn-X-Ser/Thr) | Oligosacáril transferasa (RE) | Reconocimiento celular, inmunidad, estabilidad (anticuerpos IgG) |
| O-glicosilación | Ser, Thr | Glicosiltransferasas (Golgi) | Mucinas, grupos sanguíneos ABO |
| Lipidación | Gly N-t, Cys | Aciltransferasas | Anclaje a membrana; proteínas Ras, proteínas G |
14.7. Proteólisis limitada: secuencias señal y zimógenos
14.7.1. Secuencias señal y direccionamiento
Muchas proteínas llevan incorporada en su secuencia N-terminal una secuencia señal (péptido señal) de 15–30 aminoácidos que actúa como código postal intracelular. Esta secuencia es reconocida por receptores específicos que dirigen la proteína a su compartimento de destino: retículo endoplasmático (para proteínas secretadas o de membrana), mitocondria, peroxisoma o núcleo.
Una vez que la proteína alcanza su destino, la secuencia señal es eliminada por peptidasas señal específicas. La proteína madura no la contiene. Las proteínas que contienen la secuencia señal pero aún no han sido procesadas se denominan pre-proteínas.
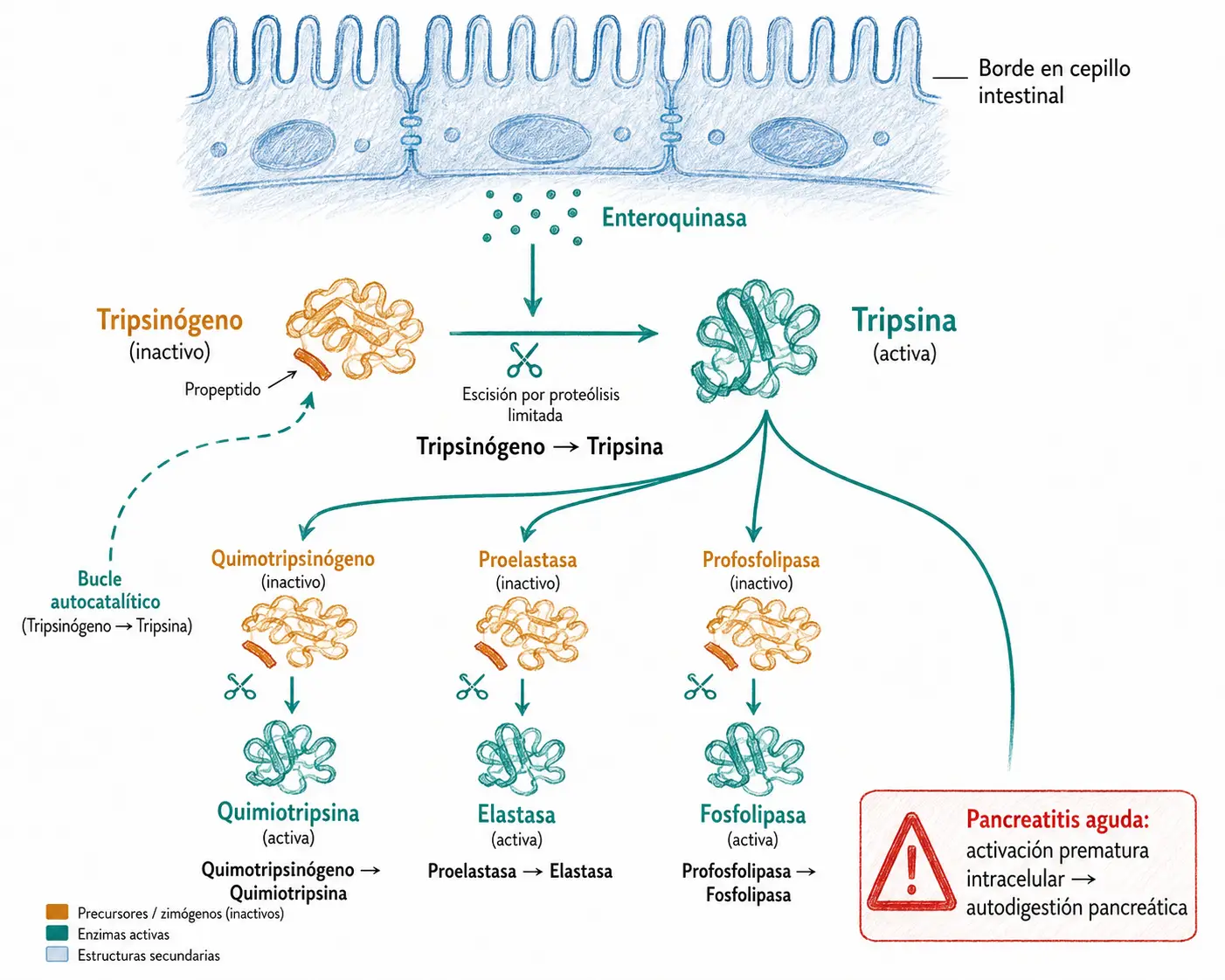
14.7.2. Zimógenos y proteólisis de activación
Los zimógenos (también llamados proenzimas) son precursores proteicos inactivos que se activan mediante proteólisis limitada: el corte preciso de uno o varios enlaces peptídicos elimina un propéptido inhibidor y reordena el sitio activo en su conformación catalítica.
Este mecanismo es especialmente importante para proteínas cuya actividad prematura sería peligrosa. Los ejemplos más relevantes son las proteasas digestivas (pepsinógeno → pepsina en el estómago por acidez; tripsinógeno → tripsina en el duodeno por enteroquinasa) y los factores de coagulación, que circulan como zimógenos y se activan en cascada solo cuando y donde se necesitan.
La pancreatitis aguda es la consecuencia de la activación prematura de los zimógenos pancreáticos dentro del propio páncreas: el tripsinógeno se convierte en tripsina intracelularmente y activa en cascada al resto de proenzimas (quimotripsinógeno, proelastasa, profosfolipasa), lo que desencadena la autodigestión del tejido pancreático.
14.8. Adición de grupos prostéticos
Muchas proteínas requieren para su función la unión covalente o no covalente de una molécula no proteica denominada grupo prostético. La síntesis de la apoenzima (parte proteica) y la disponibilidad del grupo prostético están coordinadas: cuando el grupo prostético escasea, se reduce la síntesis de la proteína correspondiente.
Grupo prostético: molécula no proteica unida covalente o no covalentemente a una proteína (apoenzima) que es imprescindible para su actividad funcional. La proteína sin su grupo prostético se denomina apoenzima; la forma activa completa es la holoenzima. Ejemplos: grupo hemo (hemoglobina), biotina (carboxilasas), FAD/FMN (deshidrogenasas flavoproteicas).
Los grupos prostéticos más relevantes en bioquímica médica son el grupo hemo (hemoglobina, mioglobina, citocromos), la biotina (carboxilasas), el FAD/FMN (deshidrogenasas flavoproteicas), el coenzima A y los metales de transición (zinc, hierro, cobre) en el centro activo de numerosas metaloproteínas.
