Este tema desarrolla las fases 2 y 3 del catabolismo presentadas en Tema 17 · Metabolismo intermediario. Se recomienda tener claros los conceptos de ΔG, ATP y carga energética antes de continuar.
16.1. Fases del metabolismo intermediario: de los principios inmediatos al acetil-CoA
El catabolismo de los tres principios inmediatos (glúcidos, lípidos y proteínas) no llega de forma directa al ciclo de Krebs. Pasa antes por una etapa intermedia en la que las moléculas complejas se reducen a un número muy pequeño de metabolitos centrales. El más importante de todos es el acetil-CoA.
Las rutas de entrada al acetil-CoA son distintas para cada principio inmediato:
- Glúcidos: la glucosa se degrada por glucólisis hasta piruvato. El piruvato entra en la mitocondria y es descarboxilado oxidativamente por el complejo piruvato deshidrogenasa (PDH) para generar acetil-CoA, CO₂ y NADH. Esta reacción es irreversible y constituye un punto de control clave: en ayuno, la PDH se inhibe para desviar el piruvato hacia la gluconeogénesis.
- Lípidos: los ácidos grasos se activan a acil-CoA en el citoplasma, entran en la mitocondria y son degradados por la β-oxidación, que libera unidades de acetil-CoA en cada vuelta del ciclo.
- Proteínas: los aminoácidos, tras la eliminación del grupo amino (transaminación o desaminación), ceden sus esqueletos carbonados como piruvato, acetil-CoA o intermediarios directos del ciclo de Krebs (oxalacetato, α-cetoglutarato, fumarato, succinato).
La glucólisis y sus mecanismos de regulación se desarrollan en detalle en T19: Glúcidos y glucólisis. La β-oxidación de ácidos grasos, en T24: Oxidación de ácidos grasos y cuerpos cetónicos. El metabolismo de aminoácidos, en T27–T29: Metabolismo de aminoácidos.
16.2. El ciclo de Krebs
16.2.1. Concepto y localización
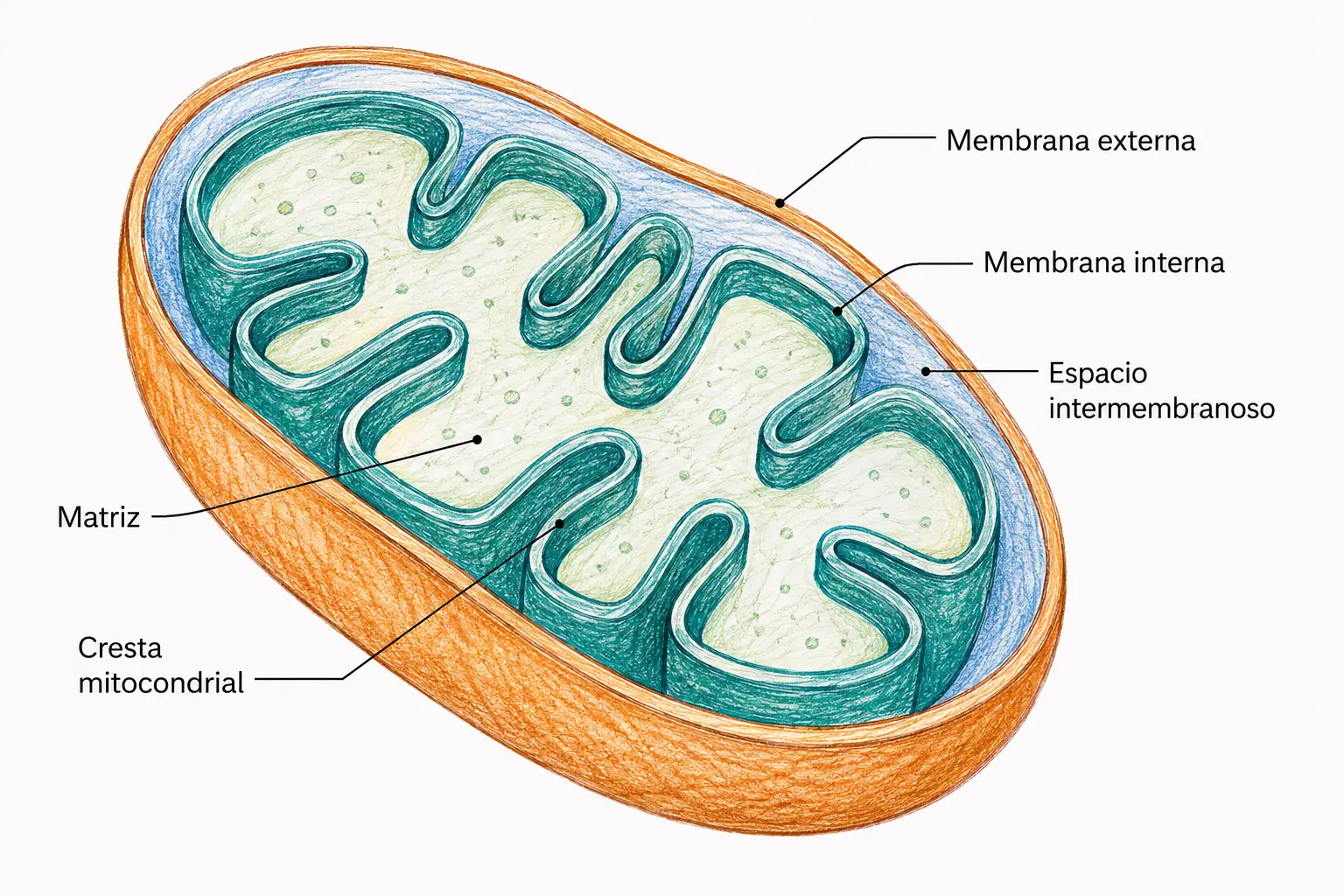
Ciclo de Krebs (ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos): conjunto de reacciones enzimáticas organizadas de forma cíclica que representa la vía final y común de la degradación oxidativa de los tres principios inmediatos. Tiene lugar íntegramente en la matriz mitocondrial.
El ciclo fue descrito por Hans Krebs en 1937, trabajo por el que recibió el Premio Nobel de Medicina en 1953. La clave del descubrimiento fue comprender que no era el piruvato directamente, sino el acetil-CoA, el sustrato real del ciclo, y que los intermediarios del ciclo actuaban como catalizadores de su propia regeneración.
El ciclo comienza cuando el acetil-CoA (2 carbonos) condensa con el oxalacetato (4 carbonos) para formar citrato (6 carbonos). A lo largo de ocho reacciones sucesivas, el citrato se transforma de nuevo en oxalacetato, liberando en el proceso los dos carbonos del acetil-CoA en forma de CO₂ y transfiriendo la energía de las oxidaciones a coenzimas reducidas (NADH y FADH₂).
16.2.2. Balance energético por vuelta
Por cada vuelta del ciclo, a partir de un acetil-CoA, se obtiene:
| Producto | Cantidad por vuelta | Destino energético |
|---|---|---|
| NADH | 3 mol | Cadena de transporte electrónico → ~2,5 ATP cada uno |
| FADH₂ | 1 mol | Cadena de transporte electrónico → ~1,5 ATP |
| GTP (o ATP) | 1 mol | Fosforilación directa a nivel de sustrato |
| CO₂ | 2 mol | Producto de desecho (eliminado por respiración) |
El rendimiento total por vuelta, contando lo que genera la cadena de transporte electrónico con los NADH y FADH₂ producidos, es de aproximadamente 10 ATP por acetil-CoA.
El ciclo de Krebs no genera ATP directamente en cantidad significativa: solo produce 1 GTP por vuelta. Su función principal es capturar la energía de oxidación en forma de coenzimas reducidas (NADH y FADH₂), que serán el combustible de la cadena de transporte electrónico.
16.2.3. Las ocho reacciones del ciclo
16.2.4. Propiedades del ciclo
El ciclo de Krebs tiene tres propiedades que lo distinguen de otras rutas metabólicas:
- Universal. Aparece en prácticamente todos los organismos aerobios. En bacterias anaerobias que carecen de él existe el ciclo del glioxalato como alternativa, pero en células eucariotas el ciclo de Krebs es la única ruta de oxidación terminal del acetil-CoA.
- Doblemente catalizado. Cada reacción tiene su enzima específica, pero además los propios metabolitos del ciclo actúan como activadores catalíticos: si se añade cualquier intermediario del ciclo al sistema, la velocidad de todo el ciclo aumenta. Esto es lo que los bioquímicos llaman «efecto catalítico de los intermediarios».
- Anfibólico. El ciclo funciona tanto en sentido catabólico (degradando acetil-CoA para generar energía) como en sentido anabólico (suministrando precursores para la síntesis de aminoácidos, nucleótidos, hemoglobina y ácidos grasos). Esta doble función exige que el ciclo esté constantemente reaprovisionado de intermediarios, lo que se consigue mediante las reacciones anapleróticas.
16.3. Reacciones anapleróticas y de secuestro
16.3.1. Reacciones anapleróticas
Si los intermediarios del ciclo de Krebs se consumen en rutas anabólicas, el ciclo se vaciaría y se detendría. Las reacciones anapleróticas («de relleno») reponen estos intermediarios para mantener el flujo del ciclo.
Reacciones anapleróticas: reacciones que reponen los intermediarios del ciclo de Krebs consumidos en rutas biosintéticas. Aseguran la continuidad del ciclo independientemente del estado anabólico de la célula.
Las más importantes son:
| Reacción | Enzima | Intermediario repuesto |
|---|---|---|
| Piruvato + CO₂ → Oxalacetato | Piruvato carboxilasa | Oxalacetato (esencial para iniciar el ciclo) |
| Piruvato → Malato | Enzima málica (NADP⁺-dependiente) | Malato |
| Aminoácidos gluconeogénicos → intermediarios del ciclo | Transaminasas + enzimas específicas | α-Cetoglutarato, succinato, fumarato, oxalacetato |
La piruvato carboxilasa tiene una importancia especial: es la enzima que garantiza que siempre haya oxalacetato disponible para condensar con el acetil-CoA en la primera reacción del ciclo. Se activa alostéricamente por acetil-CoA: cuando hay mucho acetil-CoA, la enzima se activa para producir más oxalacetato y así acelerar el ciclo.
El acetil-CoA activa alostéricamente a la piruvato carboxilasa. Cuando hay exceso de acetil-CoA, la célula lo interpreta como señal de que necesita más oxalacetato para procesar ese acetil-CoA en el ciclo. La piruvato carboxilasa responde generando ese oxalacetato. Es un ejemplo elegante de regulación por sustrato.
16.3.2. Reacciones de secuestro
La función anfibólica del ciclo implica también el proceso contrario: los intermediarios del ciclo son «secuestrados» hacia rutas biosintéticas cuando la célula los necesita:
- Oxalacetato → gluconeogénesis (vía fosfoenolpiruvato), especialmente en ayuno.
- Citrato → síntesis de ácidos grasos: el citrato sale de la mitocondria, se rompe en el citoplasma para liberar acetil-CoA citosólico, el precursor de la lipogénesis.
- α-Cetoglutarato y oxalacetato → síntesis de aminoácidos (glutamato, aspartato y sus derivados).
- Succinil-CoA → síntesis de porfirinas (hemoglobina, citocromos).
El déficit de porfirinas por alteración del succinil-CoA es el mecanismo molecular de las porfirias: enfermedades metabólicas hereditarias que cursan con fotosensibilidad cutánea, dolor abdominal y síntomas neuropsiquiátricos. La más frecuente es la porfiria aguda intermitente, por déficit de porfobilinógeno deaminasa.
16.4. Regulación del ciclo de Krebs
El ciclo de Krebs es una ruta de tal importancia que no puede regularse con un único punto de control. Se regula en múltiples niveles, de forma coordinada con el estado energético de la célula.
Los tres puntos de regulación principales son enzimas que catalizan reacciones irreversibles y responden a señales del estado redox y energético de la mitocondria:
| Enzima reguladora | Activadores | Inhibidores |
|---|---|---|
| Citrato sintetasa | Oxalacetato, ADP | ATP, NADH, succinil-CoA, citrato |
| Isocitrato deshidrogenasa | ADP, Ca²⁺ | ATP, NADH |
| α-Cetoglutarato deshidrogenasa | Ca²⁺, ADP | ATP, NADH, succinil-CoA |
El principio regulador general es coherente con la lógica energética: cuando la célula tiene exceso de ATP y NADH (carga energética alta), el ciclo se frena. Cuando hay escasez de ATP (carga energética baja), el ciclo se activa para regenerarlo. El Ca²⁺ activa las tres enzimas reguladoras en el músculo, coordinando el aumento de producción de ATP con el aumento de la actividad contráctil.
Las tres enzimas reguladoras del ciclo de Krebs responden al mismo patrón: se inhiben por los productos finales del proceso energético (ATP, NADH) y se activan cuando escasean. El Ca²⁺ funciona como señal de "demanda energética inmediata" en el músculo: cuando hay contracción, hay Ca²⁺, y el Ca²⁺ activa el ciclo para producir más ATP.
16.5. La cadena de transporte de electrones
16.5.1. Función y localización
El NADH y el FADH₂ producidos en el ciclo de Krebs (y en la glucólisis y la β-oxidación) son portadores de electrones de alta energía, pero no pueden ceder esa energía directamente al ATP. La cadena de transporte de electrones (CTE) es el sistema que extrae la energía de esos electrones de forma escalonada, en saltos controlados, acoplando cada salto a la síntesis de ATP.
La CTE está formada por cuatro complejos multiproteicos integrados en la membrana mitocondrial interna, junto con dos transportadores móviles que los conectan: la coenzima Q (ubiquinona, lipofílica, difunde por la bicapa lipídica) y el citocromo c (proteína pequeña, difunde por el espacio intermembranoso).
16.5.2. Los cuatro complejos
Los cuatro complejos forman una cadena funcional en la que los electrones fluyen siempre en el mismo sentido, desde los portadores de menor potencial de reducción hacia los de mayor potencial.
- El NADH cede sus electrones al complejo I, que los pasa a la coenzima Q.
- El FADH₂ los cede directamente al complejo II, que también los vierte sobre la coenzima Q.
- Desde aquí, el complejo III los recoge y los transfiere al citocromo c.
- El complejo IV los pasa finalmente al oxígeno, reduciéndolo a agua.
Los complejos I, III y IV acoplan el paso de electrones al bombeo de protones hacia el espacio intermembranoso, contribuyendo así al gradiente quimiosmótico.
El complejo II es una excepción. Transfiere electrones a la coenzima Q pero no bombea protones. Por eso el FADH₂, que entra en la cadena por el complejo II saltándose el primer punto de bombeo, genera menos ATP que el NADH.
La coenzima Q y el citocromo c actúan como lanzaderas móviles entre complejos: la primera dentro de la bicapa lipídica, el segundo en el espacio intermembranoso. Sin ellas, la cadena se interrumpiría porque los complejos no están en contacto directo entre sí.
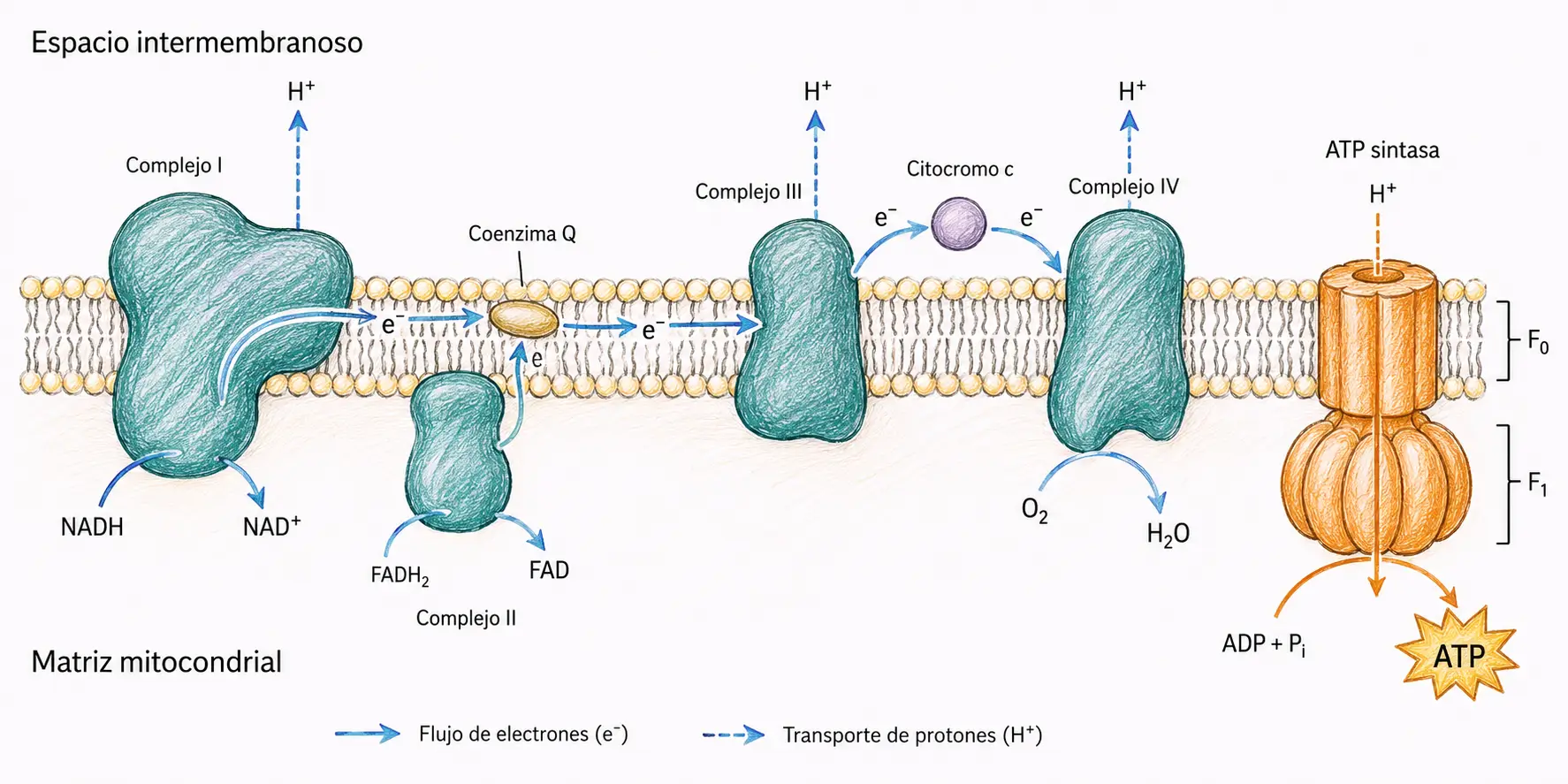
| Complejo | Nombre | Función | Bombeo de H⁺ |
|---|---|---|---|
| I | NADH-coenzima Q oxidorreductasa | Oxida NADH; transfiere los electrones a la coenzima Q | 4 H⁺ al espacio intermembranoso |
| II | Succinato-coenzima Q oxidorreductasa | Oxida FADH₂ (procedente del succinato); transfiere los electrones a la coenzima Q | No bombea H⁺ |
| III | Coenzima Q-citocromo c oxidorreductasa | Transfiere electrones de la coenzima Q al citocromo c | 4 H⁺ al espacio intermembranoso |
| IV | Citocromo c oxidasa | Transfiere electrones del citocromo c al O₂ (aceptor final), formando H₂O | 2 H⁺ al espacio intermembranoso |
El complejo II (succinato deshidrogenasa) no bombea protones y, por tanto, la oxidación del FADH₂ genera menos ATP que la del NADH. Este es el motivo por el que FADH₂ equivale a ~1,5 ATP y NADH a ~2,5 ATP en los cálculos modernos de rendimiento energético.
16.5.3. El gradiente quimiosmótico
Los complejos I, III y IV bombean protones (H⁺) desde la matriz mitocondrial al espacio intermembranoso a medida que los electrones fluyen a través de ellos. El resultado es la acumulación de protones en el espacio intermembranoso, que genera dos fuerzas simultáneas:
- Un gradiente de concentración (más H⁺ fuera que dentro).
- Un gradiente eléctrico (el espacio intermembranoso queda cargado positivamente respecto a la matriz).
Juntas, estas dos fuerzas constituyen la fuerza protón-motriz, que representa energía potencial almacenada en el gradiente electroquímico de protones. Esta energía es la que aprovecha la ATP sintasa para sintetizar ATP.
Este mecanismo, denominado teoría quimiosmótica, fue propuesto por Peter Mitchell en 1961 y le valió el Premio Nobel de Química en 1978. Su idea central era radical para la época: la energía del transporte electrónico no se almacena en un intermediario químico, sino en un gradiente iónico a través de una membrana.
Los desacoplantes son moléculas que disipan el gradiente de protones sin pasar por la ATP sintasa, por lo que la energía se libera como calor en lugar de sintetizarse ATP. El 2,4-dinitrofenol fue usado en la década de 1930 como adelgazante, con resultados fatales: la liberación incontrolada de calor causaba hipertermia, taquicardia y muerte. Hoy sigue comercializándose ilegalmente. La termogenina (UCP1) es un desacoplante fisiológico presente en el tejido adiposo pardo: su función es generar calor para la termorregulación en recién nacidos y animales en hibernación.
16.6. La fosforilación oxidativa: la ATP sintasa
La ATP sintasa (también llamada complejo V o F₀F₁-ATPasa) es la enzima que aprovecha el gradiente de protones para sintetizar ATP a partir de ADP y fosfato inorgánico. Es la principal fuente de ATP de la célula aerobia.
Está formada por dos dominios funcionales:
- F₀: dominio transmembranoso, integrado en la membrana mitocondrial interna. Forma un canal por el que fluyen los protones desde el espacio intermembranoso hacia la matriz. Este flujo de protones hace girar el dominio F₀ como un rotor molecular.
- F₁: dominio catalítico, orientado hacia la matriz. Compuesto por tres pares de subunidades α y β alternadas. El giro del rotor F₀ provoca cambios conformacionales en las subunidades β, que catalizan la síntesis de ATP mediante el mecanismo de cambio conformacional de Boyer.
La ATP sintasa es literalmente un motor molecular rotatorio. Cada 360° de rotación completa sintetiza 3 moléculas de ATP, una por cada subunidad β del dominio F₁. Para dar una vuelta completa se necesitan ~10 protones. Paul Boyer y John Walker recibieron el Premio Nobel de Química en 1997 por descubrir este mecanismo.
La síntesis de ATP por la ATP sintasa requiere que la membrana mitocondrial interna esté intacta y que el gradiente de protones se mantenga. Si la membrana se permeabiliza (por desacoplantes, toxinas o lesión celular), el gradiente colapsa y la síntesis de ATP cesa aunque el transporte electrónico siga funcionando.
16.7. Balance energético global de la oxidación completa de la glucosa
Integrando glucólisis, complejo PDH, ciclo de Krebs y fosforilación oxidativa, el balance energético de la oxidación completa de una molécula de glucosa en condiciones aerobias es:
| Etapa | NADH | FADH₂ | ATP directo | ATP total (aprox.) |
|---|---|---|---|---|
| Glucólisis | 2 | — | 2 | ~7 |
| Complejo PDH (×2) | 2 | — | — | ~5 |
| Ciclo de Krebs (×2) | 6 | 2 | 2 (GTP) | ~20 |
| Total por molécula de glucosa | ~30–32 ATP | |||
El valor clásico de 36–38 ATP por glucosa corresponde a cálculos basados en cocientes P/O teóricos (3 ATP/NADH, 2 ATP/FADH₂). Los valores modernos (~30–32 ATP) utilizan los cocientes experimentales reales (2,5 ATP/NADH, 1,5 ATP/FADH₂) y tienen en cuenta el coste energético del transporte de ADP y fosfato al interior mitocondrial. Ambos valores pueden aparecer en el examen; lo importante es entender el razonamiento.
