17.1. Definición y funciones biológicas de los glúcidos
Los glúcidos, también llamados hidratos de carbono o sacáridos, son moléculas orgánicas formadas por carbono, hidrógeno y oxígeno, en una proporción general de (CH₂O)n. El nombre hidrato de carbono refleja esta proporción pero es químicamente inexacto: los glúcidos no son carbono hidratado sino polihidroxialdehídos o polihidroxicetonas, o compuestos que los generan por hidrólisis.
Sus funciones biológicas son múltiples y no se reducen a la energía:
- Reserva energética: glucógeno en animales, almidón en vegetales.
- Combustible inmediato: la glucosa es el sustrato preferente de la mayoría de células y el obligado para el eritrocito y la neurona en condiciones normales.
- Función estructural: la celulosa forma la pared celular vegetal; la quitina, el exoesqueleto de artrópodos y la pared fúngica.
- Reconocimiento celular y señalización: los oligosacáridos de la superficie celular participan en la adhesión célula-célula, el reconocimiento inmune y la determinación del grupo sanguíneo ABO.
- Componentes de moléculas informacionales: la ribosa y la desoxirribosa son la base estructural del ARN y el ADN respectivamente.
Los glúcidos no son solo combustible. Sus funciones estructurales (celulosa, quitina), informacionales (ribosa en ARN) y de reconocimiento celular (glucoconjugados de superficie) son igualmente esenciales para la vida. Reducir los glúcidos a "fuente de energía" es uno de los errores conceptuales más frecuentes en primero de medicina.
17.2. Clasificación de los glúcidos
Los glúcidos se clasifican por su grado de polimerización:
- Monosacáridos: unidad mínima, no hidrolizable. Glucosa, fructosa, galactosa, ribosa, desoxirribosa. Se clasificación, a su vez, por dos criterios:
- Por el grupo carbonilo:
- Aldosas: el carbonilo está en C1, grupo aldehído.
- Cetosas: el carbonilo está en C2, grupo cetona.
- La glucosa es una aldosa; la fructosa, una cetosa.
- Por el número de carbonos:
- Triosas (3C), tetrosas (4C), pentosas (5C), hexosas (6C).
- Las más relevantes en metabolismo humano son las hexosas (glucosa, fructosa, galactosa) y las pentosas (ribosa, desoxirribosa).
- Por el grupo carbonilo:
- Disacáridos: dos monosacáridos unidos por un enlace glucosídico. Sacarosa, lactosa, maltosa.
- Oligosacáridos: 3–10 unidades. Frecuentes en glucoconjugados de superficie celular.
- Polisacáridos: más de 10 unidades. Almidón, glucógeno, celulosa, quitina.
Monosacárido: polihidroxialdehído o polihidroxicetona de cadena corta (3–7 carbonos) que no puede hidrolizarse a unidades más pequeñas. Es la unidad estructural básica de todos los glúcidos.
17.2. Monosacáridos
17.2.1. Estereoisomería D/L de los monosacáridos
Todos los monosacáridos con más de dos carbonos tienen al menos un carbono quiral, lo que genera isómeros ópticos. La convención D/L se basa en la configuración del carbono quiral más alejado del grupo carbonilo: si el hidroxilo de ese carbono está a la derecha en la proyección de Fischer, el monosacárido es D; si está a la izquierda, es L.
En los organismos vivos, prácticamente todos los monosacáridos son de la serie D. La glucosa fisiológica es D-glucosa. Esta especificidad es fundamental puesto que las enzimas metabólicas solo reconocen el estereoisómero D.
D/L no equivale a dextrógiro/levógiro. D indica configuración absoluta del carbono de referencia, no el sentido de rotación de la luz polarizada. La D-fructosa, por ejemplo, es levógira pese a ser de la serie D.
17.2.2. Formas cicladas: furanosa y piranosa
En solución acuosa, los monosacáridos de 5 o más carbonos no existen predominantemente en forma de cadena abierta. El grupo carbonilo reacciona con uno de los grupos hidroxilo de la propia molécula formando un hemiacetal intramolecular, por lo que la molécula se cicla.
La glucosa cicla preferentemente formando un anillo de seis miembros: la glucopiranosa. En esta ciclación, el C1 (carbonilo) se convierte en un nuevo carbono quiral denominado carbono anomérico. Según la posición del hidroxilo resultante en C1 se generan dos anómeros:
- α-D-glucopiranosa: el hidroxilo del C1 está en posición axial (abajo en la representación de Haworth).
- β-D-glucopiranosa: el hidroxilo del C1 está en posición ecuatorial (arriba).
En solución, la glucosa establece un equilibrio entre ambas formas (mutarrotación): aproximadamente 36% α y 64% β. La forma β es más estable porque el hidroxilo ecuatorial minimiza las tensiones estéricas del anillo.
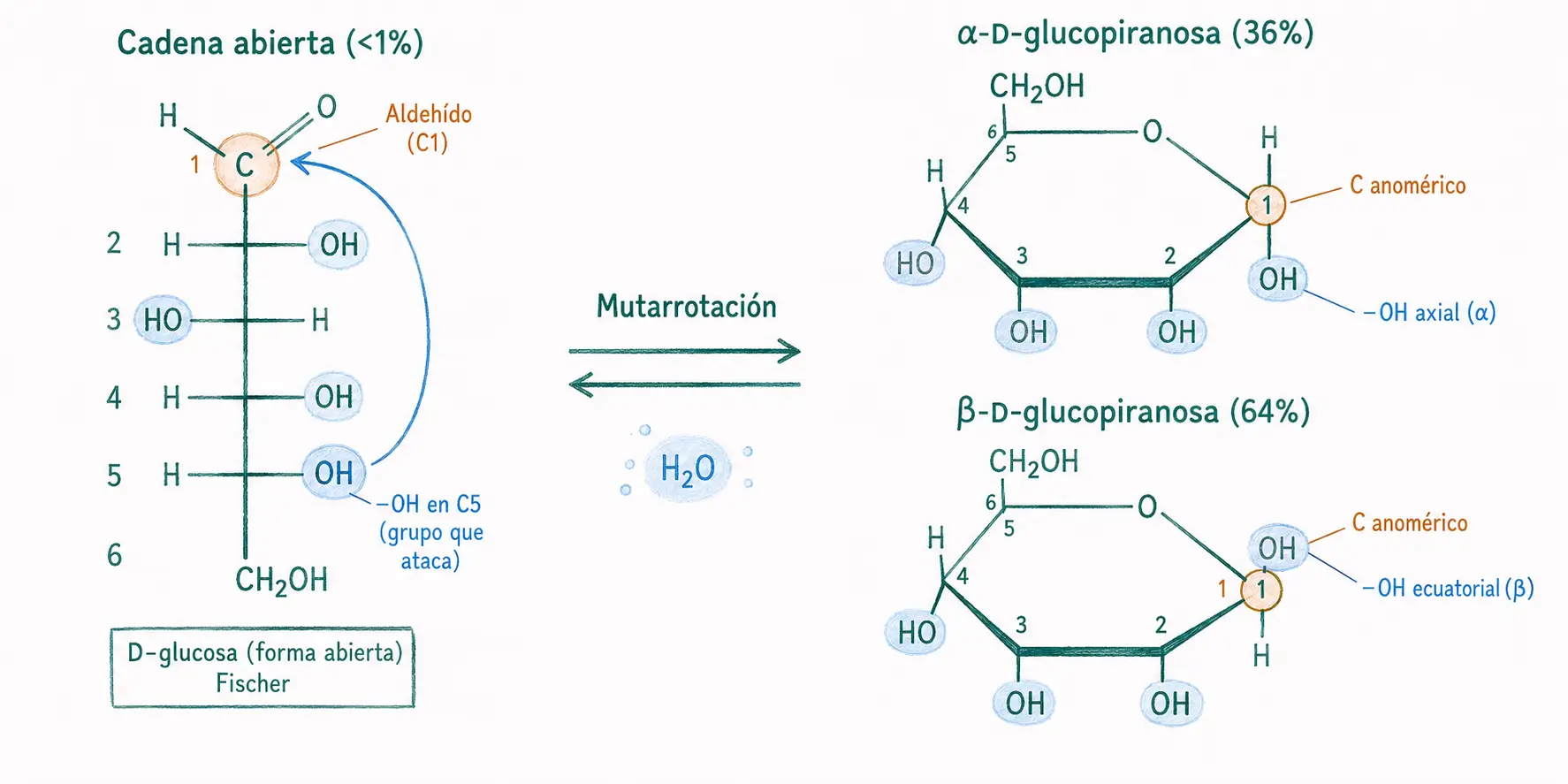
La posición del hidroxilo anomérico del C1 no es un detalle estructural menor: determina el tipo de enlace glucosídico que puede formarse y, por tanto, las propiedades del polisacárido resultante. El enlace α-1,4 da lugar al almidón (digerible); el enlace β-1,4 da lugar a la celulosa (indigerible en humanos).
17.2.3. Epímeros y azúcares reductores
Se denominan epímeros dos monosacáridos que difieren únicamente en la configuración de uno de sus carbonos quirales, sin contar el carbono anomérico. La glucosa y la galactosa son epímeras en C4: idénticas en todo excepto en la posición del hidroxilo del cuarto carbono. La glucosa y la manosa son epímeras en C2. Esta diferencia mínima tiene consecuencias enzimáticas mayores: las enzimas que metabolizan glucosa no reconocen galactosa, y viceversa.
Epímeros: monosacáridos que difieren en la configuración de un único carbono quiral (distinto del carbono anomérico). La glucosa y la galactosa son epímeras en C4; la glucosa y la manosa, en C2.
Un azúcar es reductor cuando tiene el carbono anomérico libre, es decir, sin participar en un enlace glucosídico. En esa situación el carbono anomérico puede oxidarse y reducir otros compuestos. Todos los monosacáridos son reductores. Entre los disacáridos, la lactosa y la maltosa son reductoras porque uno de sus carbonos anoméricos queda libre; la sacarosa no lo es porque el enlace glucosídico une los dos carbonos anoméricos (C1 de glucosa y C2 de fructosa), bloqueando ambos.
La detección de azúcares reductores en orina (glucosuria, galactosuria) se basa históricamente en la reacción de Benedict, que detecta cualquier azúcar reductor. Las tiras reactivas modernas son específicas de glucosa (glucosa oxidasa), por lo que no detectan galactosa ni fructosa. En un recién nacido con ictericia y glucosuria negativa en tira reactiva pero positiva en Benedict hay que descartar galactosemia.
17.2.4. Ácidos urónicos y derivados oxidados
Los monosacáridos pueden oxidarse en posiciones distintas generando derivados ácidos con funciones biológicas específicas.
El más importante es el ácido glucurónico, resultado de la oxidación del C6 de la glucosa. Su función principal es aumentar la solubilidad de moléculas hidrofóbicas. En el hígado, el ácido glucurónico se conjuga con bilirrubina, hormonas esteroideas y fármacos poco solubles para facilitar su excreción biliar o renal. Este proceso se denomina glucuronidación y es una de las reacciones de biotransformación hepática de fase II más importantes.
La glucuronidación hepática es la principal vía de eliminación de la bilirrubina. En el recién nacido, la inmadurez de la UDP-glucuronil transferasa reduce la capacidad de conjugar bilirrubina, causando la ictericia fisiológica neonatal. El fenobarbital, inductor de esta enzima, se ha usado clásicamente para acelerar la maduración de la glucuronidación en ictericias neonatales prolongadas.
17.3. Disacáridos y el enlace glucosídico
Cuando el hidroxilo anomérico de un monosacárido reacciona con un hidroxilo de otro monosacárido, se forma un enlace O-glucosídico con pérdida de agua. Este enlace es covalente, estable, y determina las propiedades del disacárido resultante.
Los tres disacáridos de mayor relevancia biológica y clínica son:
- Lactosa (galactosa β-1,4 glucosa): disacárido de la leche de mamíferos. El enlace β-1,4 requiere lactasa para su hidrólisis. El déficit de lactasa es la causa de la intolerancia a la lactosa, una de las variantes metabólicas más frecuentes en la especie humana. La lactosa es un azúcar reductor porque el C1 de la glucosa queda libre.
- Sacarosa (glucosa α-1,2 fructosa): disacárido vegetal, el azúcar de mesa. El enlace glucosídico une los dos carbonos anoméricos (C1 de glucosa y C2 de fructosa), por lo que ninguno queda libre: la sacarosa no es reductora.
- Maltosa (glucosa α-1,4 glucosa): producto de la digestión del almidón por la amilasa. Es reductora porque el C1 de una de las glucosas queda libre.
La intolerancia a la lactosa afecta a más del 70% de la población adulta mundial, con grandes variaciones geográficas. Es casi universal en Asia oriental y muy frecuente en África subsahariana, mientras que es minoritaria en poblaciones del norte de Europa donde la persistencia de lactasa es la norma evolutiva seleccionada. El diagnóstico se confirma con el test de hidrógeno espirado o la determinación genética del polimorfismo LCT-13910C>T.
17.4. Polisacáridos
Los polisacáridos son cadenas de monosacáridos unidas por enlaces glucosídicos. Pueden ser homopolisacáridos (un solo tipo de monosacárido) o heteropolisacáridos (varios tipos). Sus propiedades físicas y funcionales dependen del tipo de enlace, el grado de ramificación y la longitud de cadena.
17.4.1. Polisacáridos de reserva
- Almidón: reserva energética vegetal. Formado por dos componentes: amilosa (cadena lineal, enlaces α-1,4) y amilopectina (cadena ramificada, enlaces α-1,4 en la cadena principal y α-1,6 en los puntos de ramificación). Digerible por las α-amilasas humanas.
- Glucógeno: reserva energética animal. Estructura similar a la amilopectina pero con mayor grado de ramificación (ramificación cada 8–12 glucosas vs. cada 24–30 en la amilopectina). Su alta ramificación tiene una ventaja funcional: hay muchos extremos no reductores disponibles simultáneamente para la acción de las glucógeno fosforilasas, lo que permite una movilización muy rápida de glucosa cuando la demanda energética aumenta bruscamente.
17.4.2. Polisacáridos estructurales
- Celulosa: componente de la pared celular vegetal. Cadena lineal de glucosa con enlaces β-1,4. Las cadenas se organizan en fibrillas paralelas estabilizadas por puentes de hidrógeno, lo que le confiere una rigidez mecánica extraordinaria. Los humanos carecen de celulasas, por lo que la celulosa no es digerible, pero actúa como fibra dietética regulando el tránsito intestinal.
- Quitina: componente del exoesqueleto de artrópodos y la pared fúngica. Estructura similar a la celulosa pero con N-acetilglucosamina en lugar de glucosa. También con enlaces β-1,4.
El glucógeno está más ramificado que el almidón por una razón funcional: cuantos más extremos no reductores tenga la molécula, más rápido puede movilizarse la glucosa cuando la célula la necesita. El hígado necesita responder en segundos a una hipoglucemia: una estructura muy ramificada lo hace posible.
17.4.3. Mucopolisacáridos
Los más relevantes en medicina son:
- Ácido hialurónico: el GAG más grande del organismo. Formado por glucurónico + N-acetilglucosamina. Presente en tejido conjuntivo, líquido sinovial y humor vítreo. Da viscosidad y resistencia mecánica. Los fibroblastos lo sintetizan; la hialuronidasa lo degrada (presente en espermatozoides para penetrar el óvulo, y en algunas bacterias patógenas para abrirse paso por el tejido conjuntivo).
- Condroitín sulfato: abundante en cartílago. Formado por glucurónico + N-acetilgalactosamina sulfatada.
- Heparina: GAG de secreción producido por mastocitos. Potente anticoagulante endógeno: activa la antitrombina III, que inhibe la trombina e impide la formación de fibrina.
- Dermatán sulfato y queratán sulfato: presentes en piel, córnea y tejido conjuntivo.
Las mucopolisacaridosis son enfermedades de depósito lisosomal causadas por el déficit de enzimas que degradan los GAG. Al acumularse en los tejidos producen daño progresivo multiorgánico. Las más conocidas son el síndrome de Hurler (déficit de α-L-iduronidasa, acumulación de dermatán y heparán sulfato) y el síndrome de Hunter (déficit de iduronato-2-sulfatasa, ligado al X). Clínicamente cursan con rasgos faciales toscos, hepatoesplenomegalia, alteraciones esqueléticas y, en las formas graves, deterioro cognitivo.
La heparina es uno de los fármacos anticoagulantes más usados en medicina. Su mecanismo es la activación de la antitrombina III, que inhibe la trombina y el factor Xa. Se administra en trombosis venosa profunda, embolia pulmonar, síndrome coronario agudo y como anticoagulación en circulación extracorpórea. Su antídoto es el sulfato de protamina.
17.5. Glucoconjugados
Los glucoconjugados son moléculas formadas por glúcidos unidos covalentemente a proteínas (glucoproteínas) o lípidos (glucolípidos). Son componentes esenciales de la membrana plasmática y de la matriz extracelular.
- Glucoproteínas: los oligosacáridos se unen a residuos de serina/treonina (enlace O-glucosídico) o asparagina (enlace N-glucosídico). Los oligosacáridos de superficie forman el glucocáliz, la capa de carbohidratos que recubre la cara extracelular de la membrana. El glucocáliz participa en el reconocimiento celular, la adhesión, la protección frente a proteasas y la señalización.
- Glucolípidos: los oligosacáridos se unen al ceramida en la cara extracelular de la membrana. Los gangliósidos son glucolípidos complejos abundantes en neuronas; sus oligosacáridos participan en la señalización y en el reconocimiento célula-célula.
El sistema de grupos sanguíneos ABO está determinado por glucoconjugados de la superficie del eritrocito. Los antígenos A, B y H son oligosacáridos que difieren en el azúcar terminal añadido por glucosiltransferasas específicas. El grupo O carece de la transferasa funcional y expone el antígeno H sin modificar. La incompatibilidad ABO en transfusiones provoca hemólisis por activación del complemento mediada por anticuerpos anti-A o anti-B preformados.
17.6. Digestión y absorción de glúcidos
Los polisacáridos y disacáridos de la dieta deben hidrolizarse previamente a monosacáridos. La digestión de glúcidos comienza en la boca con la amilasa salival (ptialina) y se completa en el intestino delgado por la amilasa pancreática y las disacaridasas del borde en cepillo del enterocito (lactasa, sacarasa, maltasa).
El resultado final de la digestión es una mezcla de monosacáridos: aproximadamente 80% glucosa, 20% fructosa y galactosa. Estos monosacáridos se absorben en el enterocito mediante transportadores específicos y pasan a la circulación portal. Su transporte y captación celular (especialmente los transportadores GLUT) se desarrollan en el Tema 18 · Catabolismo de glúcidos. Glucólisis.
