9.1. Introducción: los ácidos nucleicos
9.1.1. El nucleótido: estructura y componentes
Los ácidos nucleicos son polímeros de nucleótidos. Son las moléculas que almacenan, transmiten y expresan la información genética de todos los seres vivos, y su comprensión es el punto de partida de la biología molecular.
Un nucleótido está formado por tres componentes unidos de forma covalente: una base nitrogenada, una pentosa y uno o varios grupos fosfato. La pentosa actúa como elemento central: se une a la base por su carbono 1′ mediante un enlace N-glucosídico, y al fosfato por su carbono 5′ mediante un enlace éster.
Las bases nitrogenadas se clasifican en dos grupos según su estructura. Las purinas (adenina y guanina) tienen una estructura bicíclica de dos anillos fusionados. Las pirimidinas (citosina, timina y uracilo) tienen un único anillo. Esta diferencia de tamaño es relevante para el apareamiento: en la doble hélice siempre se enfrenta una purina con una pirimidina, lo que mantiene un diámetro uniforme a lo largo de toda la molécula.
Cuando la base se une a la pentosa sin el grupo fosfato, la molécula resultante se denomina nucleósido. Al añadir uno o más fosfatos al carbono 5′ del nucleósido se obtiene el nucleótido. Los nucleótidos trifosfato, como el ATP o el GTP, son además las unidades de activación energética que utiliza la célula para polimerizar los ácidos nucleicos y para transferir energía en el metabolismo.
9.1.2. Diferencias entre ADN y ARN
El ADN y el ARN comparten la arquitectura básica de nucleótidos encadenados por enlaces fosfodiéster, pero difieren en tres aspectos estructurales con consecuencias funcionales importantes.
- La primera diferencia es la pentosa. El ADN contiene 2′-desoxirribosa, que carece del grupo hidroxilo en el carbono 2′. El ARN contiene ribosa, con ese grupo OH presente. Este detalle aparentemente menor hace al ARN químicamente más reactivo e inestable: el grupo 2′-OH puede atacar el enlace fosfodiéster adyacente y fragmentar la cadena. La mayor estabilidad del ADN lo hace idóneo como depósito permanente de información genética.
- La segunda diferencia es una de las bases pirimidínicas. El ADN utiliza timina (5-metilúracilo), mientras que el ARN utiliza uracilo, que carece del grupo metilo en posición 5. Funcionalmente son equivalentes como bases apareantes con adenina, pero la timina del ADN permite distinguir las citosinas desaminadas espontáneamente (que se convierten en uracilo) y repararlas sin ambigüedad.
- La tercera diferencia es la estructura de cadena. El ADN se presenta habitualmente como doble hélice. El ARN es monocatenario, aunque se pliega sobre sí mismo formando estructuras secundarias locales mediante apareamiento intramolecular de bases complementarias.
| Característica | ADN | ARN |
|---|---|---|
| Pentosa | 2'-desoxirribosa (sin OH en C2') | Ribosa (OH en C2') |
| Base pirimidínica diferencial | Timina (5-metilúracilo) | Uracilo (sin grupo metilo en C5) |
| Estructura de cadena | Bicatenario (doble hélice) | Monocatenario (con plegamiento intramolecular local) |
| Estabilidad química | Alta: ausencia del 2'-OH impide la autocatálisis | Menor: el 2'-OH puede atacar el enlace fosfodiéster |
| Función principal | Almacenamiento permanente de información genética | Intermediario, adaptador, catalizador y regulador |
9.1.3. La cadena polinucleotídica: polaridad y enlace fosfodiéster
Los nucleótidos se unen entre sí mediante enlaces fosfodiéster: el grupo fosfato del nucleótido entrante se une al carbono 3′ del nucleótido anterior, formando un puente que conecta el carbono 3′ de una pentosa con el carbono 5′ de la siguiente.
Esta química de unión imprime una polaridad a la cadena. Un extremo termina con un fosfato libre unido al carbono 5′ de la primera pentosa (extremo 5′) y el otro extremo termina con un grupo hidroxilo libre en el carbono 3′ de la última pentosa (extremo 3′). Esta direccionalidad no es trivial: todas las polimerasas que sintetizan ácidos nucleicos, tanto en replicación como en transcripción, añaden nucleótidos exclusivamente en el extremo 3′, de modo que la síntesis avanza siempre en sentido 5’→3′.
En la doble hélice del ADN, las dos cadenas son antiparalelas: una corre en sentido 5’→3′ y la opuesta en sentido 3’→5′. Este antiparalelismo es una consecuencia geométrica obligada del apareamiento de bases de Watson-Crick: solo con las cadenas orientadas en sentidos contrarios los pares A=T y G≡C adoptan la geometría correcta para formar sus puentes de hidrógeno.
La cadena polinucleotídica tiene polaridad 5'→3'. Las dos hebras de la doble hélice son antiparalelas y complementarias: donde una tiene A la otra tiene T, y donde una tiene G la otra tiene C.
9.1.4. Otras funciones de los nucleótidos
Los nucleótidos no son únicamente las unidades de construcción de los ácidos nucleicos. En su forma libre, con dos o tres grupos fosfato, actúan como moléculas funcionales con papeles centrales en el metabolismo celular.
Nucleótido libre: nucleótido que no está incorporado a una cadena polinucleotídica y actúa de forma autónoma como molécula funcional. Su actividad depende del número de grupos fosfato, del tipo de base y de las modificaciones que pueda llevar.
La función más conocida es la transferencia de energía:
- El ATP (adenosín trifosfato) y el GTP (guanosín trifosfato) son las principales monedas energéticas de la célula.
- La hidrólisis del enlace entre el segundo y el tercer fosfato libera energía utilizable para impulsar reacciones termodinámicamente desfavorables, contracciones musculares, transporte activo y síntesis de macromoléculas.
- El propio proceso de polimerización de los ácidos nucleicos consume nucleótidos trifosfato: al incorporarse a la cadena creciente, se libera pirofosfato, cuya hidrólisis proporciona la energía que desplaza el equilibrio hacia la síntesis.
Los nucleótidos forman parte de coenzimas imprescindibles para el metabolismo intermediario.
- El NAD⁺ y el NADP⁺ contienen adenina y actúan como transportadores de electrones en la glucólisis, el ciclo de Krebs y la cadena respiratoria.
- El FAD, derivado de riboflavina pero con adenosín monofosfato en su estructura, actúa como aceptor de electrones en reacciones de oxidación.
- La coenzima A (CoA) contiene también un nucleótido de adenina y es el transportador universal de grupos acilo en el metabolismo de lípidos, carbohidratos y aminoácidos.
Los nucleótidos cíclicos actúan como segundos mensajeros en la transducción de señales.
- El AMP cíclico (AMPc) se genera a partir de ATP por la adenilato ciclasa cuando determinadas hormonas se unen a sus receptores de membrana.
- El AMPc activa la proteína cinasa A (PKA), que fosforila proteínas diana y amplifica la señal hormonal.
- El GMP cíclico (GMPc) opera de forma análoga en otras vías de señalización, entre ellas la del óxido nítrico y la fototransducción retiniana.
Los análogos sintéticos de nucleótidos son fármacos que interfieren en la replicación o la transcripción al incorporarse a la cadena de ADN o ARN en lugar del nucleótido natural. Al carecer del grupo 3'-OH necesario para continuar la polimerización, actúan como terminadores de cadena. El aciclovir (análogo de guanosina) inhibe la ADN polimerasa del virus herpes. El AZT (zidovudina, análogo de timidina) inhibe la transcriptasa inversa del VIH. La gemcitabina (análogo de citidina) y la 5-fluorouracilo se usan en quimioterapia por su capacidad de bloquear la replicación en células tumorales de alta proliferación. El mecanismo de acción de estos fármacos se basa directamente en la química del enlace fosfodiéster descrita en 9.1.3.
9.2. El ácido desoxirribonucleico (ADN)
9.2.1. Fuerzas que estabilizan la estructura secundaria del ADN
La estructura secundaria del ADN se estabiliza por la acción conjunta de tres tipos de fuerzas.
- El apilamiento hidrofóbico (stacking) entre bases adyacentes de la misma cadena es la fuerza estabilizadora dominante. Los anillos aromáticos de las bases se apilan en paralelo, minimizando el contacto con el agua. La suma cooperativa de estas interacciones a lo largo de toda la molécula genera una estabilidad muy superior a la de cualquier enlace individual.
- Los puentes de hidrógeno entre las bases enfrentadas de las dos cadenas aportan especificidad al apareamiento: A=T forma dos puentes y G≡C forma tres. Son la razón por la que la Tm es proporcional al contenido en G+C, pero no son la fuerza estabilizadora principal.
- Las interacciones electrostáticas de los grupos fosfato con el agua y con cationes divalentes (especialmente Mg²⁺) estabilizan el esqueleto azúcar-fosfato en el entorno acuoso intracelular.
Las tres fuerzas actúan simultáneamente y de forma cooperativa: la suma de miles de interacciones débiles a lo largo de toda la molécula genera una estabilidad muy superior a la de cualquier enlace aislado. Es precisamente esta cooperatividad la que hace que la desnaturalización del ADN tenga un comportamiento de transición abrupta en torno a la Tm, en lugar de producirse de forma gradual.
La disposición antiparalela de las cadenas y el ángulo de unión entre pares de bases generan dos cavidades longitudinales en la superficie de la hélice: el surco mayor y el surco menor. El surco mayor es más ancho y profundo, y expone más información química de las bases hacia el exterior. La mayoría de las proteínas que reconocen secuencias específicas de ADN (factores de transcripción, enzimas de restricción) lo hacen a través del surco mayor. El surco menor también es accesible, pero ofrece menos especificidad de reconocimiento.
El apilamiento hidrofóbico entre bases adyacentes es la fuerza estabilizadora dominante de la doble hélice, no los puentes de hidrógeno. Estos últimos aportan especificidad de apareamiento, pero menos energía total.
Afirmar que los puentes de hidrógeno son la fuerza principal que mantiene la doble hélice es un error habitual. El apilamiento hidrofóbico aporta más energía de estabilización.
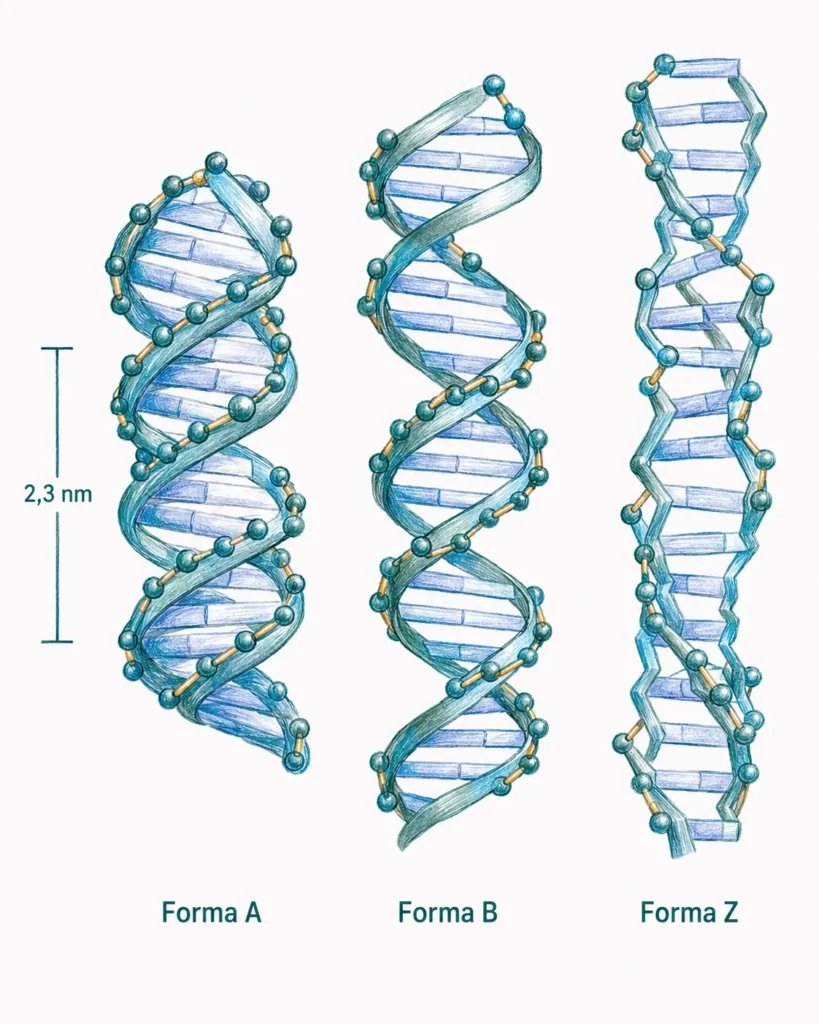
9.2.2. Conformaciones del ADN: formas B, A y Z
El ADN no adopta una única estructura rígida. Según la secuencia, el estado de hidratación y las condiciones del entorno, la doble hélice puede asumir al menos tres conformaciones bien caracterizadas.
La forma B es la conformación predominante en las condiciones fisiológicas normales y la descrita originalmente por Watson y Crick en 1953. Es dextrógira, con un diámetro de 20 Å y aproximadamente diez pares de bases por vuelta helicoidal. La distancia entre bases adyacentes es de 3,4 Å. Los surcos mayor y menor son claramente distinguibles; el surco mayor es más ancho y profundo, y es el principal punto de reconocimiento por proteínas reguladoras.
La forma A también es dextrógira, pero más corta y ancha que la forma B: tiene unos once pares de bases por vuelta y un diámetro mayor. Las bases están más inclinadas respecto al eje de la hélice. Esta conformación aparece en condiciones de baja hidratación y, de forma relevante, es la adoptada por los híbridos ADN-ARN y por el ARN de doble cadena. No es la forma habitual del ADN genómico in vivo.
La forma Z es levógira y se caracteriza porque el esqueleto fosfato dibuja un trayecto en zigzag, de ahí su nombre. Aparece en regiones con alta repetición de dinucleótidos CpG y se favorece por la metilación del ADN. La metilación de citosinas en contexto CpG es un mecanismo epigenético de silenciamiento: los genes en regiones Z-metiladas tienden a no transcribirse. Es la única forma levógira conocida in vivo.
La metilación del ADN en promotores de genes supresores de tumores (como BRCA1 o MLH1) los silencia sin alterar la secuencia. Esta inactivación epigenética es un mecanismo frecuente en la carcinogénesis. A diferencia de una mutación, las marcas de metilación son potencialmente reversibles, lo que las convierte en diana de fármacos epigenéticos (inhibidores de ADN metiltransferasa, como azacitidina).
9.2.3. Estructuras no canónicas: cruciforme, triple hélice y G-quadruplex
El ADN de triple hélice se forma cuando una tercera cadena se introduce en el surco mayor de una doble hélice y establece enlaces con las bases ya apareadas. Para ello no se separan completamente las dos hebras originales: la tercera cadena reconoce la cara externa de los pares de bases ya formados mediante apareamientos de Hoogsteen, una geometría de enlace de hidrógeno distinta a la de Watson-Crick.
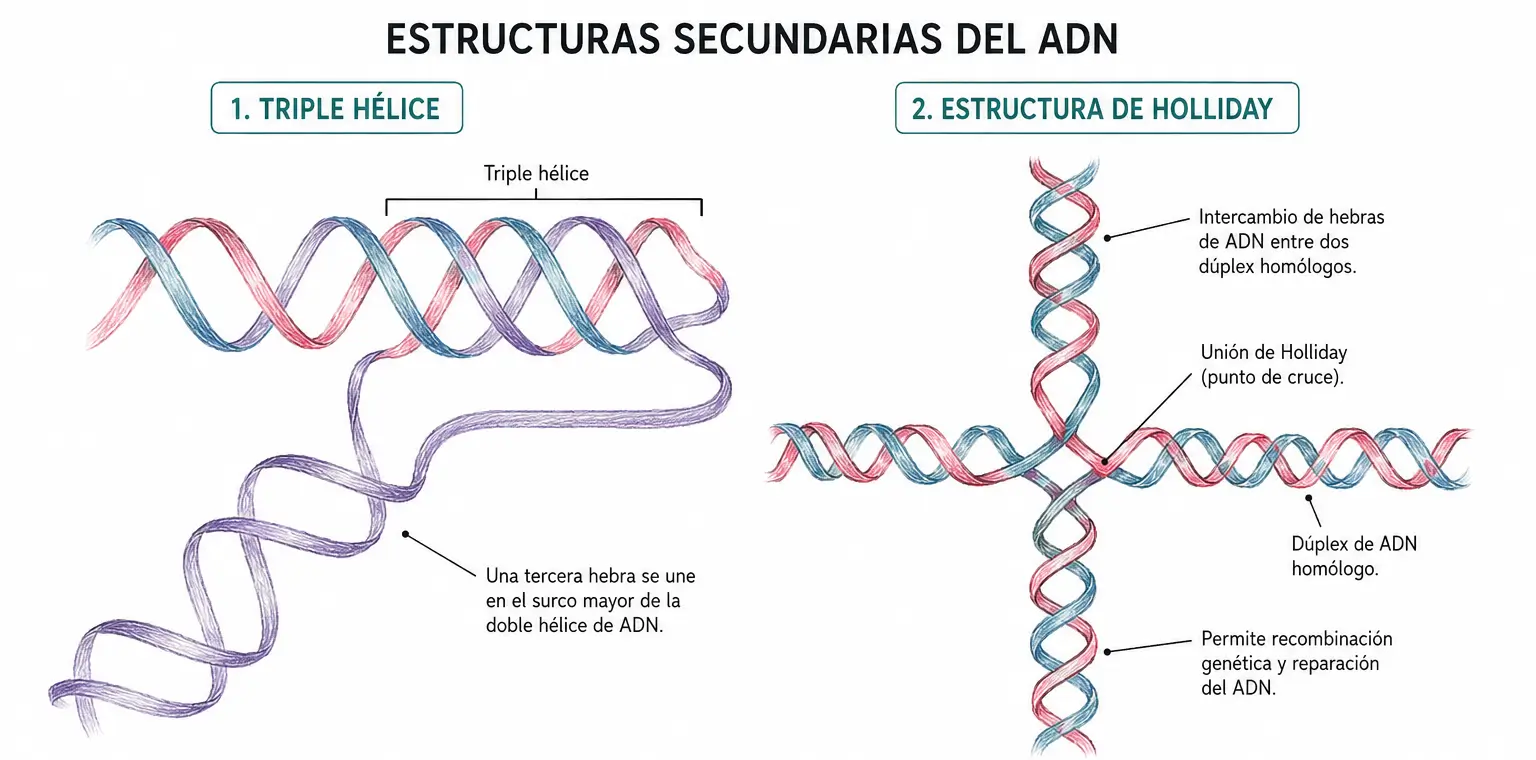
La cadena entrante puede ser de secuencia similar a una de las hebras o complementaria a ella, dependiendo de las condiciones. Las tripletas de bases resultantes son combinaciones como T·A=T o C·G≡C.
El ADN de triple hélice es una estructura de interés en biología molecular, pero su relevancia fisiológica in vivo sigue siendo objeto de investigación. Se estudia como herramienta potencial para el silenciamiento dirigido de genes específicos.
Estructuras cruciformes. Aparecen en secuencias con repeticiones palindrómicas invertidas (inverted repeats): cada cadena se pliega sobre sí misma formando una horquilla, y las dos horquillas generan una estructura en cruz. Se han descrito en regiones reguladoras del genoma. Un ejemplo biológico bien caracterizado es la unión de Holliday, una estructura cruciforme transitoria que aparece durante la recombinación homóloga entre dos dúplex de ADN homólogos.
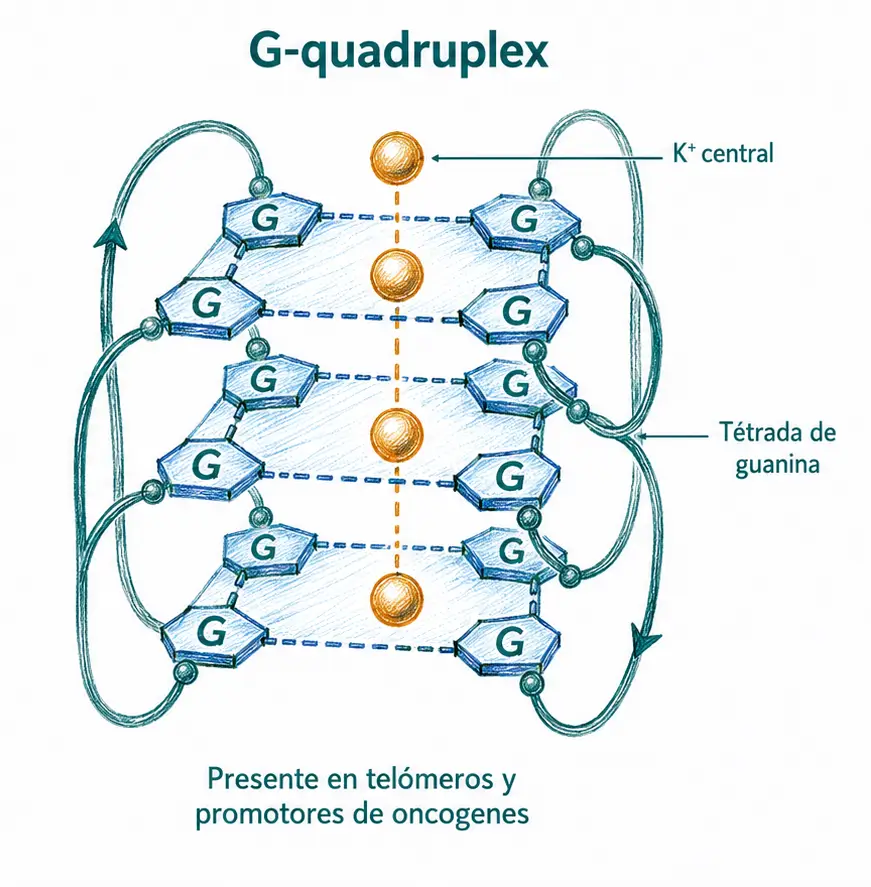
G-quadruplex (tetraplex G). Se forman en secuencias ricas en guanina mediante tétradas de cuatro guaninas estabilizadas por un catión monovalente central (K⁺ o Na⁺). Abundan en los telómeros y en los promotores de proto-oncogenes como c-MYC o KRAS. Los apareamientos responsables se denominan de Hoogsteen, distintos de los de Watson-Crick de la doble hélice. Los G-quadruplex son una diana terapéutica activa en oncología.
El ADN de triple hélice requiere la separación local de las dos hebras para integrar una tercera cadena. Dos cadenas son iguales y una es complementaria. Es una estructura de interés en biología molecular, pero su relevancia fisiológica in vivo sigue siendo objeto de investigación.
9.2.4. Desnaturalización del ADN
La desnaturalización del ADN es la pérdida de su estructura secundaria por rotura de las fuerzas que mantienen unidas las dos hebras, principalmente el apilamiento hidrofóbico y los puentes de hidrógeno entre bases. El resultado es la separación de las dos cadenas polinucleotídicas, que quedan como hebras simples. La desnaturalización no rompe los enlaces fosfodiéster, por lo que la secuencia de cada cadena se conserva íntegramente.
Puede producirse por calor, pH extremo o agentes desnaturalizantes químicos.
En condiciones adecuadas es reversible: al eliminar el agente desnaturalizante y enfriar lentamente, las cadenas complementarias se reaparan espontáneamente restaurando la doble hélice. Este proceso se denomina renaturalización o hibridación, y tiene numerosas aplicaciones en biología molecular y diagnóstico.
El efecto hipercrómico es el aumento de absorbancia a 260 nm que se produce al desnaturalizar el ADN. En la doble hélice, el apilamiento de bases reduce su capacidad de absorber luz ultravioleta (hipocromicidad). Al separarse las cadenas, las bases quedan libres y recuperan su absorbancia máxima. Este fenómeno permite monitorizar la desnaturalización espectrofotométricamente.
Tm (temperatura de fusión o temperatura de desnaturalización): temperatura en el que el 50% del ADN está desnaturalizado. Relacionada directamente con el contenido en guanina y citosina del ADN.
La desnaturalización reversible del ADN es la base de técnicas como la hibridación, el Southern blotting o la PCR. Estas aplicaciones se desarrollan en el tema de técnicas de biología molecular.
9.2.5. Compactación del ADN
En procariotas, el ADN cromosómico es circular y su longitud es varios órdenes de magnitud mayor que el diámetro celular. Para que quepa en el nucleoide, el ADN se compacta mediante superenrollamiento: las dos hebras de la doble hélice se enrollan adicionalmente sobre el propio eje de la molécula, generando tensión torsional. En condiciones fisiológicas el ADN bacteriano se encuentra en superenrollamiento negativo, lo que facilita la separación local de las hebras durante la replicación y la transcripción.
Las moléculas de ADN que tienen la misma secuencia pero difieren en su grado de superenrollamiento se denominan topoisómeros. El equilibrio entre topoisómeros está regulado por las topoisomerasas, enzimas que modifican la topología del ADN cortando y religando hebras. Las topoisomerasas de tipo I cortan una sola hebra; las de tipo II cortan las dos hebras simultáneamente, pasando un segmento de ADN a través del corte antes de religarlo.
Las topoisomerasas regulan el estado de superenrollamiento: las de tipo I cortan una hebra y las de tipo II cortan las dos. Las fluoroquinolonas (ciprofloxacino) inhiben la topoisomerasa II bacteriana (girasa) y son antibióticos de uso habitual. Los inhibidores de topoisomerasa II eucariota (etopósido, doxorrubicina) se usan en quimioterapia.
Las fluoroquinolonas bloquean la ADN girasa (topoisomerasa II bacteriana), impidiendo la replicación del ADN bacteriano. Este mecanismo explica su actividad antibiótica de amplio espectro. Los fármacos quimioterápicos como el etopósido actúan sobre la topoisomerasa II eucariota, bloqueando la replicación de las células tumorales.
En eucariotas, los ~2 metros de ADN haploide deben compactarse hasta caber en un núcleo de 6-10 µm. Esto requiere varios niveles sucesivos de empaquetamiento.
El primer nivel es el nucleosoma: unos 147 pares de bases de ADN se enrollan 1,7 vueltas alrededor de un octámero de histonas formado por dos copias de cada una de las histonas H2A, H2B, H3 y H4. El resultado visible al microscopio electrónico es la imagen de «collar de cuentas» (fibra de 11 nm).
La histona H1 no forma parte del octámero. Se une al ADN enlazador, el segmento de entre 20 y 80 pares de bases que conecta nucleosomas consecutivos, y favorece la compactación de los nucleosomas en una fibra de orden superior de 30 nm.
Los niveles superiores de compactación implican la formación de bucles de cromatina anclados a la matriz nuclear, estructuras radiales y, finalmente, el cromosoma metafásico visible al microscopio óptico (diámetro ~700 nm).
La compactación no es uniforme ni estática. La heterocromatina es cromatina muy compactada y transcripcionalmente silenciosa; la eucromatina está menos compactada y es accesible a la maquinaria de transcripción.
Las modificaciones covalentes de las histonas (acetilación, metilación, fosforilación, ubiquitinación) alteran la afinidad del ADN por el octámero y el reclutamiento de proteínas reguladoras. La acetilación de lisinas en H3 y H4 neutraliza su carga positiva, afloja el contacto con el ADN y favorece la transcripción. La combinación de estas marcas constituye el llamado código de histonas, parte del lenguaje epigenético de la célula.
La organización del cromosoma metafásico, la morfología según la posición del centrómero y los cromosomas especiales (plumoso, politénico) se desarrollan en el cromosoma metafásico en Biología.
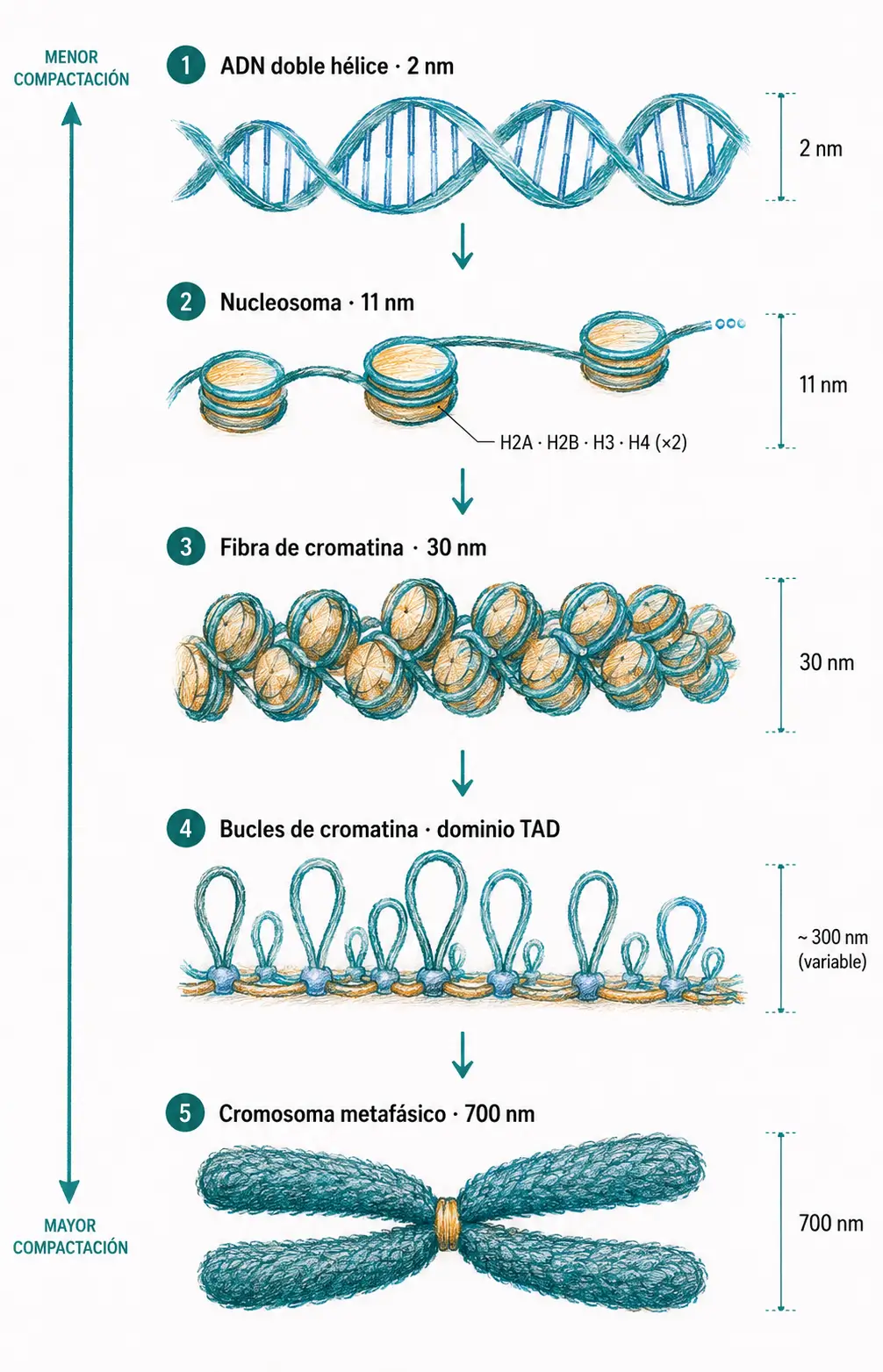
El nucleosoma está formado por un octámero de histonas H2A, H2B, H3 y H4 (dos copias de cada una) más ~147 pb de ADN. La histona H1 actúa sobre el ADN enlazador y no pertenece al octámero central.
9.2.6. Organización del genoma: secuencias únicas, repetidas y el valor C
El genoma eucariota no es una colección homogénea de genes. La mayor parte del ADN no codifica proteínas, pero las secuencias no codificantes tienen funciones estructurales, reguladoras y evolutivas.
Un fenómeno llamado valor C paradójico ilustra bien esta realidad: no existe correlación directa entre el tamaño del genoma y la complejidad del organismo. Algunos anfibios y plantas tienen genomas mucho mayores que el humano. Lo que diferencia a los organismos no es la cantidad de ADN sino la organización y regulación de la información que contiene.
En los procariotas, el cromosoma está muy compactado y prácticamente toda su secuencia es funcional. Casi todo el ADN se transcribe y la mayor parte de lo transcrito se traduce. En los eucariotas, en cambio, los genes aparecen dispersos a lo largo de cada cromosoma, separados por largas extensiones de ADN no codificante, y su organización interna es más compleja.
Las secuencias del ADN eucariota se clasifican en tres grandes categorías según su número de copias.
- Las secuencias únicas o de copia única representan aproximadamente el 70% del genoma. Están presentes una sola vez por genoma haploide. Incluyen los genes que codifican proteínas (con sus intrones y exones, que en conjunto representan solo un 3% del ADN total), los pseudogenes (copias inactivas de genes funcionales que han perdido la capacidad de expresarse) y extensas regiones de ADN espaciador de función no completamente caracterizada.
- Las secuencias moderadamente repetidas representan aproximadamente el 20% del genoma y están presentes cientos de veces. Incluyen genes que requieren tasas de síntesis muy elevadas: los genes de histonas, los que codifican ARNr y los que codifican ARNt. La existencia de múltiples copias garantiza que la demanda celular pueda satisfacerse.
- Las secuencias altamente repetidas están presentes miles de veces. Dentro de ellas se distinguen dos grupos:
- El ADN satélite está formado por secuencias muy cortas (3-10 nucleótidos) repetidas en tándem, concentradas en los centrómeros y telómeros, donde cumple funciones estructurales esenciales para la segregación cromosómica y la protección de los extremos.
- El ADN repetitivo disperso se distribuye por todo el genoma e incluye las secuencias Alu (aproximadamente 300 nucleótidos, repetidas cientos de miles de veces, derivadas de retrotransposición) y los transposones, elementos genéticos móviles capaces de migrar de una región a otra del genoma mediante mecanismos de corte-empalme o de retrotransposición.
Retrotransposición: mecanismo por el cual un elemento genético se copia a sí mismo en forma de ARN, que luego se retrotranscribe a ADN e se inserta en una nueva posición del genoma. Es el mecanismo responsable de la amplificación de las secuencias Alu y de los elementos LINE en el genoma humano.
La expansión patológica de secuencias repetidas en tándem dentro de genes funcionales causa enfermedades por repetición de trinucleótidos.
El síndrome del X frágil (repetición CGG en FMR1), la enfermedad de Huntington (CAG en HTT) y la distrofia miotónica (CTG en DMPK) son los ejemplos más relevantes. En todos los casos, el número de repeticiones supera un umbral que altera la expresión o la función de la proteína codificada.
El mecanismo molecular de estas enfermedades se desarrolla en el tema de variación genética.
La organización del genoma humano (número de cromosomas, distribución génica, secuencias repetidas y su nomenclatura) se desarrolla con mayor detalle desde la perspectiva de la biología celular en Tema 23 · El genoma humano.
9.2.7. Estructura de un gen: región reguladora, exones e intrones
El concepto de gen ha evolucionado. En sentido estricto, un gen no es simplemente un fragmento de ADN que codifica una proteína: es la unidad funcional mínima capaz de producir una molécula de ARN de forma regulada.
Gen: unidad funcional de información genética que incluye la región transcrita y las secuencias reguladoras necesarias para controlar su expresión. Un gen puede codificar una proteína o un ARN funcional (ARNr, ARNt, miARN). Sin región reguladora no puede hablarse de gen en sentido molecular estricto.
Todo gen eucariota consta de dos componentes:
- La región reguladora controla cuándo, dónde y en qué cantidad se transcribe el gen. Puede estar inmediatamente adyacente a la región transcrita o a miles de nucleótidos de distancia.
- El elemento regulador más próximo es el promotor, la secuencia donde se ensambla el complejo de iniciación de la transcripción.
- Los elementos reguladores distales (potenciadores, silenciadores) actúan sobre el promotor por contacto físico, posible gracias al plegamiento tridimensional de la cromatina.
- La región transcrita es la que la ARN polimerasa copia en ARN. Se divide en exones e intrones.
- Los exones son los segmentos que se conservan en el ARN mensajero maduro y que en último término codifican la proteína o el ARN funcional.
- Los intrones son secuencias intercaladas entre los exones que se transcriben formando parte del pre-ARNm pero se eliminan durante el procesamiento post-transcripcional. Cuantitativamente, los intrones constituyen la mayor parte de la región transcrita: la proporción intrón/exón es inversa a lo intuitivo.
Los intrones constituyen la mayor parte del gen estructural, no los exones. Esta aparente ineficiencia tiene ventajas evolutivas: el splicing alternativo permite generar múltiples proteínas diferentes a partir de un mismo gen, y la separación de exones en módulos facilita la evolución de nuevas combinaciones de dominios proteicos.
El ARN polimerasa transcribe tanto exones como intrones en un único transcrito continuo llamado pre-ARNm o ARN heterogéneo nuclear (ARNhn). Este transcrito primario sufre tres modificaciones antes de exportarse al citoplasma: adición de un cap en el extremo 5′, eliminación de los intrones por splicing y adición de una cola poli-A en el extremo 3′. El resultado es el ARNm maduro, listo para ser traducido.
El splicing alternativo, los mecanismos de control post-transcripcional y los ARN no codificantes se desarrollan en el tema 12 de este bloque. El proceso de transcripción y la maquinaria de la ARN polimerasa se tratan en el Tema 11 · Transcripción (síntesis de ARN).
La arquitectura del gen eucariota se introduce desde la perspectiva de la biología celular en Tema 23 · El genoma humano
9.3. El ácido ribonucleico (ARN)
9.3.1. Tipos y funciones del ARN
Todos los ARN son monocatenarios, aunque muchos presentan regiones de doble cadena por plegamiento intramolecular. Esta capacidad de plegarse sobre sí mismos les permite adoptar estructuras secundarias y terciarias complejas, con funciones tanto estructurales como catalíticas.
En la siguiente tabla se desglosan los tipos de ARN más destacados.
| Tipo | Abreviatura | Abundancia | Presente en | Función principal |
|---|---|---|---|---|
| ARNr ARN ribosómico | ARNr | ~80 % | Procariotas y eucariotas | Componente estructural y catalítico del ribosoma; actividad peptidiltransferasa |
| ARNt ARN de transferencia | ARNt | ~15 % | Procariotas y eucariotas | Transporta aminoácidos activados al ribosoma; adaptador entre codón y aminoácido |
| ARNm ARN mensajero | ARNm | 4–5 % | Procariotas y eucariotas | Plantilla para la síntesis de proteínas; heterogéneo y de vida media corta |
| ARNhn ARN heterogéneo nuclear | ARNhn | Variable | Eucariotas | Precursor nuclear del ARNm; incluye intrones antes del procesamiento |
| ARNsn ARN pequeño nuclear | ARNsn | Minoritario | Eucariotas | Maduración del ARNhn: eliminación de intrones (splicing) |
| ARNsc ARN pequeño citoplasmático | ARNsc | Minoritario | Eucariotas | Dirige proteínas recién sintetizadas hacia sus destinos celulares |
| ARNv ARN viral | ARNv | — | Virus ARN | Genoma viral; puede actuar directamente como ARNm o requerir transcripción previa |
| ncRNA ARN no codificantes reguladores | miRNA, siRNA, lncRNA | Variable | Eucariotas | Control postranscripcional de la expresión génica → ver T12 |
En las últimas décadas se han descrito numerosas clases de ARN no codificantes con funciones reguladoras: los microARN (miRNA) y los ARN de interferencia pequeños (siRNA) silencian genes a nivel postranscripcional, y los ARN largos no codificantes (lncRNA) participan en la regulación epigenética. Algunos son ya la base de fármacos aprobados. Su mecanismo de acción se desarrolla en el tema de control postranscripcional de la expresión génica.
9.3.2. Estructura del ARN de transferencia
El ARNt es el adaptador molecular que conecta el código genético con la síntesis de proteínas: por un extremo reconoce el codón del ARNm y por el otro transporta el aminoácido correspondiente.
Su estructura secundaria característica es el trébol: cuatro regiones de doble cadena (tallos) separadas por cuatro bucles. El bucle anticodón contiene el triplete de bases que reconoce el codón del ARNm por complementariedad. El extremo 3′-CCA es el punto de carga del aminoácido; esta secuencia es idéntica en todos los ARNt. El bucle D y el bucle TΨC participan en el reconocimiento por las aminoacil-ARNt sintetasas, las enzimas que cargan el aminoácido correcto.
En tres dimensiones, el trébol se pliega en una forma de L que aproxima el bucle anticodón y el extremo aceptor a los dos sitios funcionales del ribosoma.
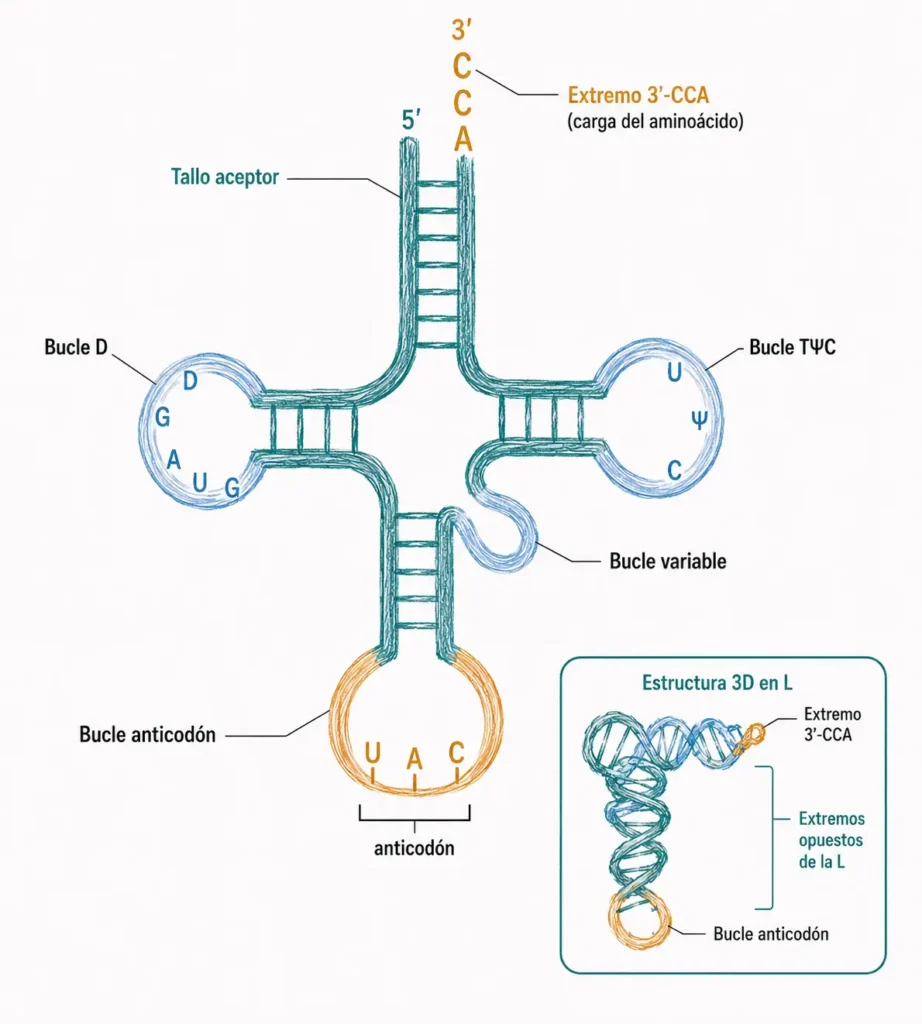
El ARNt tiene estructura secundaria en trébol con cuatro bucles. El bucle anticodón reconoce el ARNm; el extremo 3'-CCA es el punto de carga del aminoácido. Todos los ARNt comparten esta arquitectura conservada.
9.3.3. El ARN ribosómico y la actividad catalítica del ARN
El ARNr es el componente mayoritario del ribosoma y no tiene función meramente estructural: la actividad peptidiltransferasa, que cataliza la formación del enlace peptídico, reside en el ARNr 23S (procariotas) o 28S (eucariotas), no en las proteínas ribosómicas. El ribosoma es, por tanto, una ribozima, un ARN con actividad catalítica.
La existencia de ribozimas apoya la hipótesis del mundo de ARN: en el origen de la vida, el ARN habría actuado simultáneamente como molécula informacional y catalítica, antes de que la división de funciones entre ADN (almacén) y proteínas (catalizadores) se estableciera.
Los antibióticos que inhiben la síntesis de proteínas (aminoglucósidos, macrólidos, tetraciclinas, cloranfenicol) actúan sobre subunidades del ribosoma bacteriano, aprovechando las diferencias entre el ARNr procariota (70S) y eucariota (80S). Esto se desarrolla en el tema de traducción.
