13.1. El núcleo en interfase: visión general
El núcleo es el orgánulo más voluminoso y visible de la célula eucariota. Contiene el material genético y coordina todos los eventos relacionados con su metabolismo: replicación, transcripción y reparación del ADN.
El núcleo tiene un comportamiento cíclico: existe como estructura definida durante la interfase, pero desaparece durante la división celular, cuando la envoltura nuclear se fragmenta y la cromatina se condensa en cromosomas. Al finalizar la división, el núcleo se reconstituye en las células hijas. Por esta razón, cuando hablamos de núcleo siempre nos referimos al núcleo interfásico.
La función principal del núcleo es albergar y proteger el ADN del citoplasma (donde enzimas lisosomales, el citoesqueleto y otras moléculas podrían degradarlo o dañarlo), manteniendo al mismo tiempo un tráfico intenso de información con el citoplasma a través de los complejos de poro.
13.2. Características generales del núcleo
13.2.1. Forma
La forma del núcleo se relaciona con la forma de la célula. Las células esféricas tienen núcleos redondeados; las células fusiformes o cilíndricas, núcleos ovalados.
En condiciones normales los núcleos son regulares, aunque existen excepciones fisiológicas como los leucocitos polimorfonucleares, con núcleos multilobulados, o el espermatozoide, con núcleo en forma de pera.
Las células tumorales presentan núcleos muy irregulares con formas aberrantes, siendo este uno de los criterios morfológicos de malignidad en anatomía patológica.
13.2.2. Tamaño
El núcleo mide habitualmente entre 5 y 8 μm de diámetro y es visible al microscopio óptico. Su tamaño es proporcional al de la célula. La relación núcleo-citoplasma se mantiene relativamente constante a lo largo de la vida celular.
Cuando el núcleo aumenta excesivamente de volumen respecto al citoplasma, la célula se divide para restablecer el equilibrio. Las células tumorales tienen núcleos desproporcionadamente grandes: otro criterio de malignidad.
13.2.3. Número y ubicación
La mayoría de células eucariotas son mononucleadas (tienen un solo núcleo). Algunas células son binucleadas (ej. hepatocitos) o polinucleadas (ej. osteoclastos, células musculares esqueléticas).
Las células polinucleadas tienen dos orígenes posibles: la división del núcleo sin división del citoplasma (cariocinesis sin citocinesis, como en los plasmodios) o la fusión de células mononucleadas formando sincitios (como en el músculo esquelético).
La ubicación del núcleo dentro de la célula es característica de cada tipo celular — central en los hepatocitos, excéntrico en las células secretoras.
13.3. Composición química del núcleo
El núcleo contiene cuatro tipos principales de moléculas:
- ADN: asociado a proteínas formando la cromatina. Nunca existe como ADN desnudo en la célula viva.
- Proteínas: el componente más abundante en masa. Se clasifican en histonas (proteínas básicas que se unen al ADN formando los nucleosomas) y no histonas (proteínas con función estructural, enzimática o reguladora como polimerasas, topoisomerasas, factores de transcripción, laminas).
- ARN: en tránsito entre el núcleo y el citoplasma. Incluye ARNm, ARNr en proceso de maduración, ARNt, y diversas especies de ARN no codificante implicadas en procesamiento y regulación.
- Sales inorgánicas: en pequeña proporción, contribuyen al mantenimiento del pH y la estructura iónica del núcleo.
La tinción de Feulgen tiñe específicamente el ADN de color rosa-magenta y es estequiométrica, es decir, permite cuantificar la cantidad de ADN por núcleo y determinar su ploidía. Se usa en histopatología para identificar células con contenido anormal de ADN, como las tumorales.
13.4. La envoltura nuclear
La envoltura nuclear es el sistema de membranas que separa el contenido nuclear del citoplasma. Está formada por dos membranas lipídicas concéntricas:
- La membrana nuclear interna está en contacto con el contenido nuclear y contacta con la lámina nuclear densa en su cara nucleoplásmica. Ambas membranas tienen composición proteica distinta a pesar de su continuidad.
- La membrana nuclear externa está en contacto con el citoplasma y tiene ribosomas adheridos en su cara citosólica. Está conectada físicamente con las cisternas del retículo endoplásmico rugoso, con el que es funcionalmente continua.
Las dos membranas están separadas por un espacio perinuclear de ~20-40 nm y se fusionan en los puntos donde se insertan los complejos de poro nuclear.
13.4.1. La lámina nuclear densa
La lámina nuclear densa es una red proteica que tapiza la cara interna de la membrana nuclear interna. Constituye el citoesqueleto del núcleo y mantiene la forma y la integridad estructural del núcleo. También sirve de anclaje para la cromatina y los complejos de poro.
Está formada por proteínas de filamento intermedio denominadas laminas, que se ensamblan formando una red cuadrangular. Se distinguen dos grupos:
- Laminas de tipo A (lamina A y lamina C): codificadas por el mismo gen (LMNA), con diferente procesamiento. La lamina A es un filamento más largo que la lamina C. Contactan con la cromatina y participan en su posicionamiento espacial dentro del núcleo.
- Laminas de tipo B (lamina B1 y B2): codificadas por genes distintos. Contactan principalmente con la membrana nuclear interna a través de proteínas transmembrana específicas: LAP1, LAP2, MAN, emerina y LBR (receptor de lamina B).
| Tipo | Gen | Contacto principal | Función |
|---|---|---|---|
| Lamina A | LMNA (mismo gen que C, procesamiento alternativo) | Cromatina y lamina C | Posicionamiento de la cromatina · soporte estructural |
| Lamina C | LMNA (mismo gen que A, filamento más corto) | Cromatina y lamina A | Posicionamiento de la cromatina · soporte estructural |
| Lamina B1 | LMNB1 (gen independiente) | Membrana nuclear interna (vía LAP, emerina, LBR) | Anclaje de la lámina a la membrana interna |
| Lamina B2 | LMNB2 (gen independiente) | Membrana nuclear interna (vía LAP, emerina, LBR) | Anclaje de la lámina a la membrana interna |
Al inicio de la mitosis, las laminas se fosforilan por las CDK mitóticas. Esto desestabiliza la red de la lámina densa, provoca el desmontaje del complejo de poro y desencadena la desaparición de la envoltura nuclear. Al final de la mitosis, la defosforilación de las laminas permite la reconstitución de la envoltura en las células hijas.
Las mutaciones en el gen LMNA producen un grupo de enfermedades denominadas laminopatías, entre las que se incluyen la progeria de Hutchinson-Gilford (envejecimiento prematuro acelerado), distintas formas de distrofia muscular de Emery-Dreifuss y algunas cardiomiopatías dilatadas. La alteración de la lámina nuclear compromete la integridad mecánica del núcleo y el posicionamiento de la cromatina, con consecuencias devastadoras especialmente en tejidos sometidos a estrés mecánico como el músculo y el corazón.
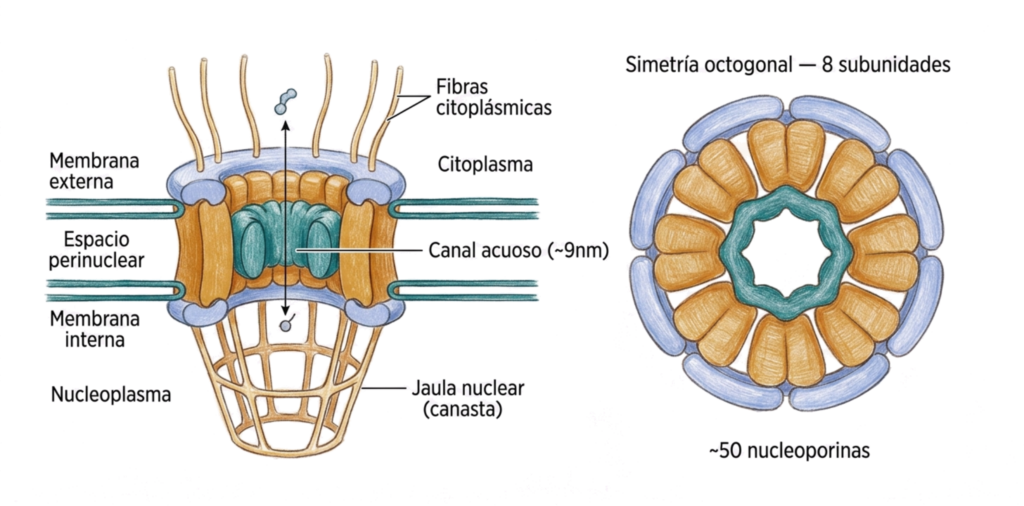
13.5. El complejo de poro nuclear
13.5.1. Estructura: nucleoporinas
Cada complejo de poro está formado por unas 50 proteínas distintas denominadas nucleoporinas, que se ensamblan formando una estructura con simetría octogonal: ocho unidades repetidas alrededor del eje central.
Las nucleoporinas se organizan en tres subunidades principales:
- Subunidad columnar. Está anclada a la envoltura nuclear mediante subunidades luminares (glucoproteínas en el espacio perinuclear). Forma la estructura de soporte del complejo.
- Subunidad anular. Está localizada en el centro del poro, responsable de su apertura o cierre. Regula el acceso al canal central.
- Subunidad del anillo. De cada unidad parten fibras proteicas que se proyectan hacia el citoplasma y hacia el núcleo. En la parte nuclear, estas fibras se unen formando la jaula nuclear o canasta nuclear, una estructura en forma de cesta que participa en el procesamiento y exportación del ARNm.
13.5.2. Transporte núcleo-citoplasma
El transporte a través del complejo de poro es bidireccional y altamente selectivo.
Existen dos modalidades de transporte:
- Difusión pasiva: el canal central del poro es acuoso, con ~9 nm de ancho y ~15 nm de largo. Por este canal pasan libremente, sin gasto energético, moléculas menores de ~5.000 Da (p. ej. iones, metabolitos pequeños, algunas proteínas muy pequeñas).
- Transporte activo: las moléculas grandes (proteínas nucleares, subunidades ribosomales, ARN) requieren transporte activo con gasto de GTP. Para ello necesitan:
- Las proteínas que deben importarse al núcleo poseen señales de localización nuclear (NLS). Las NLS son secuencias cortas de 4-8 aminoácidos generalmente cargados positivamente (Arg, Lys, Pro) que son reconocidas por receptores de importación. Estas señales pueden estar enmascaradas por proteínas inhibidoras o por fosforilación cuando la proteína no debe entrar. Solo se exponen cuando el núcleo las requiere.
- Las moléculas que deben exportarse del núcleo (ARNm, ARNt, subunidades ribosomales) poseen señales de exportación nuclear (NES) reconocidas por receptores de exportación.
| Característica | Difusión pasiva | Transporte activo |
|---|---|---|
| Tamaño de la molécula | < 5.000 Da | > 5.000 Da (proteínas grandes, ARN, subunidades ribosomales) |
| Gasto energético | No — difusión a favor de gradiente | Sí — hidrólisis de GTP (sistema Ran) |
| Señal requerida | No | Sí — NLS (importación) o NES (exportación) |
| Receptor necesario | No | Sí — carioferinas (importinas o exportinas) |
| Velocidad | Rápida | Más lenta — proceso regulado |
| Ejemplos | Iones, metabolitos, agua | Histonas, ARN polimerasa, ARNm, subunidades ribosomales |
Los receptores de importación y exportación nuclear se denominan carioferinas (importinas y exportinas respectivamente) y pertenecen a la misma familia génica. Reconocen la señal de la carga, se unen a las nucleoporinas y facilitan el paso a través del poro.
13.5.3. El sistema Ran-GTP/GDP
La direccionalidad del transporte activo está determinada por el gradiente de la proteína Ran entre el núcleo y el citoplasma:
- En el núcleo predomina Ran-GTP, mantenido por la proteína Ran-GEF (guanine exchange factor), localizada en el nucleoplasma, que convierte Ran-GDP en Ran-GTP.
- En el citoplasma predomina Ran-GDP, mantenido por la proteína Ran-GAP (GTPase-activating protein), localizada en el citoplasma, que estimula la hidrólisis de GTP a GDP.
Este gradiente determina el comportamiento de las carioferinas:
- En el citoplasma (Ran-GDP): las importinas se unen a la carga con NLS y la transportan hacia el núcleo. Las exportinas liberan su carga al disociarse de Ran-GDP.
- En el núcleo (Ran-GTP): las importinas liberan su carga al unirse a Ran-GTP. Las exportinas se unen a la carga con NES en presencia de Ran-GTP y la transportan hacia el citoplasma.
El gradiente Ran-GTP/Ran-GDP es el reloj molecular que orienta el transporte: define qué es "dentro" y qué es "fuera" del núcleo. Ran-GTP = núcleo = liberar importaciones y cargar exportaciones. Ran-GDP = citoplasma = cargar importaciones y liberar exportaciones.
13.6. La cromatina
La cromatina es el complejo de ADN y proteínas que constituye el material genético en el núcleo interfásico. Cromatina y cromosoma son la misma sustancia en dos estados de condensación diferentes: la cromatina es el estado interfásico, disperso y funcional; el cromosoma es el estado condensado que aparece durante la división celular.
13.6.1. Eucromatina y heterocromatina
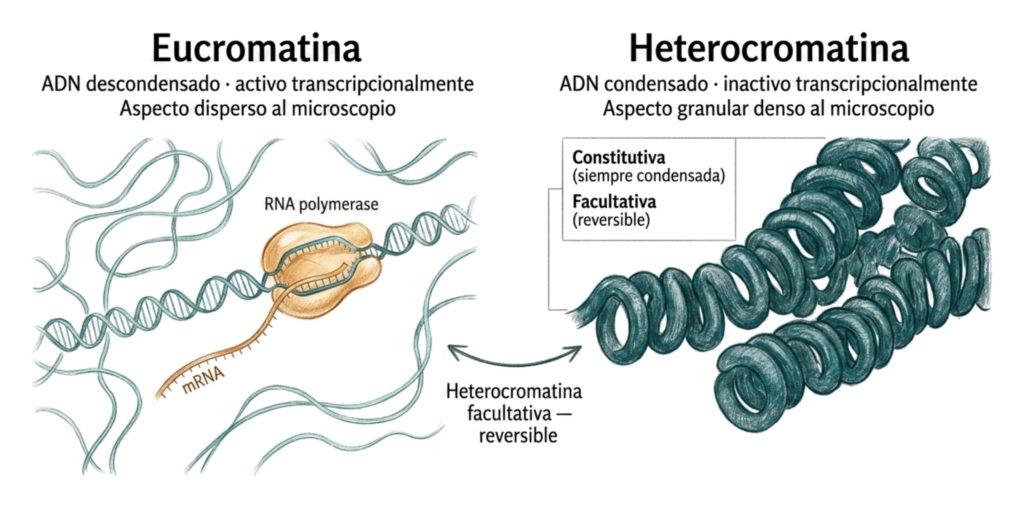
Al microscopio óptico se distinguen dos estados de la cromatina según su grado de condensación:
- La eucromatina aparece como un granulado fino y disperso. Corresponde a regiones del genoma activas transcripcionalmente. El ADN está descondensado y accesible a la maquinaria de transcripción. La mayor parte de la cromatina en una célula activa es eucromatina.
- La heterocromatina aparece como gránulos gruesos y oscuros, son regiones densamente condensadas. Corresponde a ADN inactivo transcripcionalmente. Se distinguen dos tipos:
- Heterocromatina constitutiva: siempre está condensada, en todas las células y en todos los momentos del ciclo celular. Corresponde a regiones con pocas o ninguna secuencia codificante. Corresponde con centrómeros, telómeros y el brazo largo del cromosoma Y. Su condensación permanente es estructural, no reguladora.
- Heterocromatina facultativa: puede condensarse o descondensarse en función del estado de activación génica. El ejemplo más conocido es el corpúsculo de Barr, uno de los dos cromosomas X en las células femeninas se inactiva y condensa formando heterocromatina facultativa visible al microscopio óptico.
13.6.2. Organización de la cromatina en el núcleo
La cromatina no se distribuye aleatoriamente en el núcleo interfásico. Se distinguen cuatro localizaciones características:
- Cromatina condensada perinuclear: heterocromatina adherida a la cara interna de la envoltura nuclear, en contacto con la lámina densa.
- Cromatina condensada perinucleolar: heterocromatina que rodea al nucleolo.
- Gránulos de heterocromatina dispersos: islas de cromatina condensada distribuidas por el nucleoplasma.
- Área intercromatínica: constituida por eucromatina laxa, en transcripción activa. Al microscopio electrónico aparece como un granulado fino de color gris claro.
