7.1. Concepto de señalización intracelular
La comunicación intercelular no termina cuando un mediador se une a su receptor. Esa unión es solo el primer paso. Para que la célula responda, la información debe transmitirse al interior y convertirse en cambios funcionales.
La señalización intracelular es el proceso por el que la activación de un receptor produce una respuesta dentro de la célula. Esta respuesta puede consistir en activar una enzima, abrir un canal iónico, modificar el metabolismo, inducir secreción o cambiar la expresión de genes.
En los receptores de superficie, el ligando no entra en la célula. La señal se transmite mediante cambios conformacionales del receptor y mediante proteínas intracelulares asociadas.
Proceso por el que la activación de un receptor se transforma en una respuesta dentro de la célula mediante proteínas transductoras, enzimas efectoras, segundos mensajeros y cambios en la actividad proteica o génica.
7.2. Elementos básicos de una vía de señalización intracelular
En una vía de señalización pueden distinguirse varios elementos funcionales.
- El primer mensajero es la molécula extracelular que inicia la señal. Puede ser una hormona, un neurotransmisor, una citocina, un factor de crecimiento o un mediador paracrino.
- El receptor reconoce al primer mensajero y cambia de conformación. Ese cambio permite transmitir la información hacia el interior celular.
- La proteína transductora conecta el receptor con una proteína efectora. En este tema, las más importantes son las proteínas G
- El efector suele ser una enzima o un canal iónico. Su activación produce cambios intracelulares.
- El segundo mensajero es una molécula intracelular que aparece, aumenta o se moviliza después de la activación del receptor. Su función es transmitir y amplificar la señal dentro de la célula.
| Elemento | Localización | Función | Ejemplos |
|---|---|---|---|
| Primer mensajero | Extracelular | Inicia la señal al unirse al receptor | Adrenalina · glucagón · acetilcolina · histamina |
| Receptor | Membrana plasmática o interior celular | Reconoce el ligando y activa la vía | Receptor β-adrenérgico · receptor muscarínico |
| Transductor | Cara interna de la membrana | Transmite la señal hacia el efector | Proteínas G |
| Efector | Membrana o citoplasma | Genera segundos mensajeros o modifica canales | Adenilato ciclasa · fosfolipasa C |
| Segundo mensajero | Intracelular | Amplifica y distribuye la señal | AMPc · GMPc · IP3 · DAG · Ca²⁺ |
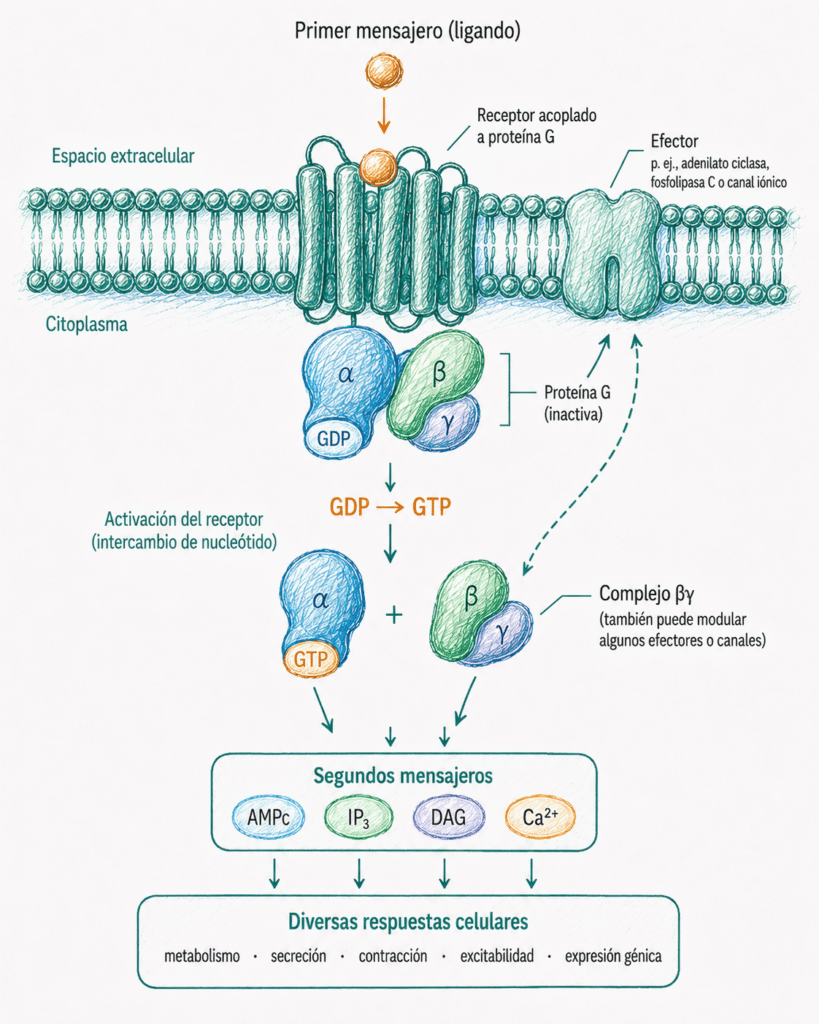
El primer mensajero actúa fuera de la célula. El segundo mensajero actúa dentro de la célula.
7.3. Receptores acoplados a proteínas G
Los receptores acoplados a proteínas G son una de las familias más importantes de receptores de superficie. Participan en la respuesta a hormonas, neurotransmisores, mediadores inflamatorios y estímulos sensoriales.
Tienen una estructura característica: una única cadena proteica atraviesa la membrana plasmática siete veces, formando siete segmentos transmembrana. Por eso también se denominan receptores de siete dominios transmembrana.
Estos receptores no tienen actividad enzimática propia. Su función es reconocer un ligando extracelular y activar una proteína G situada en la cara interna de la membrana plasmática.
Un mismo mediador puede producir respuestas distintas según el receptor al que se una. Esto explica, por ejemplo, que la adrenalina pueda tener efectos diferentes en vasos, corazón, bronquios o tejido adiposo.
Receptor acoplado a proteína G
Receptor de superficie con siete segmentos transmembrana que, tras unirse a su ligando, activa una proteína G heterotrimérica.
7.4. Proteínas G heterotriméricas
Las proteínas G asociadas a estos receptores se llaman heterotriméricas porque están formadas por tres subunidades: α, β y γ.
La subunidad α es la que une GDP o GTP. Cuando está unida a GDP, la proteína G está inactiva. Cuando está unida a GTP, se activa y puede regular efectores intracelulares.
Las subunidades β y γ permanecen habitualmente unidas formando el complejo βγ. También pueden participar en la regulación de algunos canales iónicos y otras proteínas.
En reposo, la proteína G se encuentra asociada a la cara interna de la membrana y puede interactuar con el receptor. La subunidad α permanece unida a GDP. Cuando el ligando activa el receptor, la subunidad α intercambia GDP por GTP y se separa funcionalmente del complejo βγ.
7.4.1. Ciclo de activación de las proteínas G
El funcionamiento de una proteína G puede entenderse como un interruptor molecular.
En reposo, la subunidad α está unida a GDP. Cuando el ligando activa el receptor, el receptor facilita que la subunidad α libere GDP y una GTP.
La unión de GTP activa la subunidad α. Esta subunidad α-GTP puede separarse del complejo βγ y actuar sobre enzimas efectoras, como la adenilato ciclasa o la fosfolipasa C.
La señal se apaga porque la subunidad α tiene actividad GTPasa. Es decir, puede hidrolizar el GTP a GDP y fosfato inorgánico. Cuando vuelve a estar unida a GDP, se inactiva y se reasocia con βγ.
| Fase | Estado molecular | Consecuencia funcional |
|---|---|---|
| Reposo | α-GDP asociada a βγ | Proteína G inactiva |
| Activación del receptor | El ligando se une al receptor | El receptor cambia de conformación |
| Intercambio de nucleótido | GDP se sustituye por GTP en la subunidad α | La proteína G se activa |
| Separación funcional | α-GTP se separa de βγ | Se regulan enzimas efectoras o canales iónicos |
| Inactivación | α hidroliza GTP a GDP + Pi | La señal se apaga |
| Reasociación | α-GDP vuelve a unirse a βγ | Se recupera el estado basal |
La proteína G está activa cuando la subunidad α está unida a GTP e inactiva cuando está unida a GDP.
7.4.2. Tipos funcionales de proteínas G: Gs, Gi y Gq
No todas las proteínas G activan la misma vía. La respuesta depende del tipo de subunidad α.
- La proteína Gs estimula la adenilato ciclasa. Como consecuencia, aumenta la concentración de AMPc.
- La proteína Gi inhibe la adenilato ciclasa. Por tanto, disminuye la concentración de AMPc.
- La proteína Gq activa la fosfolipasa C. Esta vía genera IP3 y DAG, dos segundos mensajeros derivados de fosfolípidos de membrana.
| Proteína G | Efector principal | Segundo mensajero | Resultado general |
|---|---|---|---|
| Gs | Adenilato ciclasa | ↑ AMPc | Activación de PKA |
| Gi | Adenilato ciclasa | ↓ AMPc | Disminución de PKA |
| Gq | Fosfolipasa C | ↑ IP3 · ↑ DAG · ↑ Ca²⁺ | Activación de PKC y vías dependientes de Ca²⁺ |
Los receptores adrenérgicos son un buen ejemplo de diversidad funcional. Los receptores β suelen acoplarse a Gs, mientras que muchos receptores α1 se acoplan a Gq y los α2 a Gi. Por eso la adrenalina puede producir respuestas distintas según el receptor expresado por cada tejido.
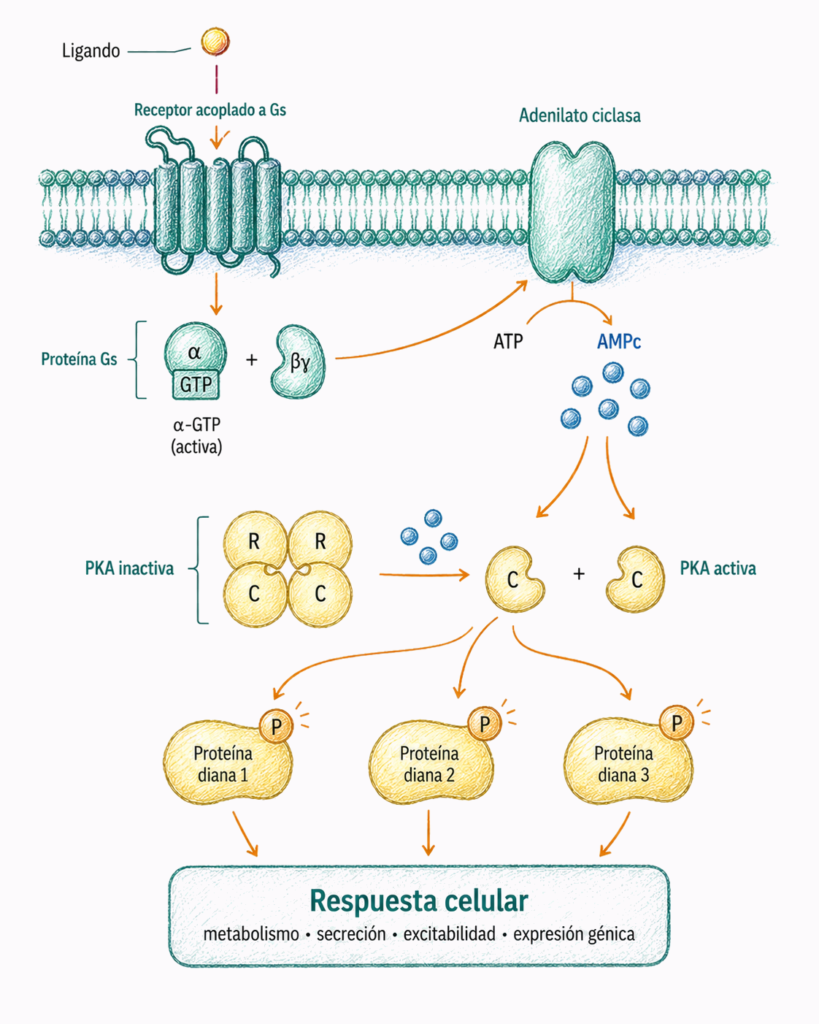
7.5. Vía adenilato ciclasa — AMPc — proteína quinasa A
La adenilato ciclasa es una enzima de membrana que cataliza la formación de AMPc a partir de ATP.
Cuando un receptor activa una proteína Gs, la subunidad αs-GTP estimula la adenilato ciclasa. Esto aumenta la concentración intracelular de AMPc.
El AMPc actúa como segundo mensajero. Su principal función clásica es activar la proteína quinasa A o PKA.
La PKA fosforila proteínas en residuos de serina y treonina. Esta fosforilación modifica la actividad de proteínas diana, que pueden ser enzimas metabólicas, canales iónicos, proteínas contráctiles o factores de transcripción.
La vía AMPc-PKA participa en procesos como metabolismo energético, secreción endocrina, excitabilidad celular y respuesta a hormonas.
Por tanto, el AMPc no suele actuar directamente sobre la respuesta final, sino que activa proteínas intermedias capaces de modificar muchas dianas celulares.
| Paso | Molécula | Función |
|---|---|---|
| 1 | Ligando | Activa un receptor acoplado a Gs |
| 2 | Gs | Activa la adenilato ciclasa |
| 3 | Adenilato ciclasa | Convierte ATP en AMPc |
| 4 | AMPc | Activa PKA |
| 5 | PKA | Fosforila proteínas diana |
7.5.1. Fosfodiesterasas y apagado de la señal por AMPc
La señal mediada por AMPc debe apagarse cuando deja de ser necesaria. El mecanismo principal es la acción de las fosfodiesterasas.
Las fosfodiesterasas degradan el AMPc y lo convierten en AMP. Al disminuir el AMPc, se reduce la activación de PKA.
Además, las proteínas fosforiladas por PKA pueden ser desfosforiladas por fosfatasas. Esto permite revertir la respuesta y recuperar el estado basal.
Este equilibrio entre quinasas y fosfatasas es fundamental para que la señalización celular sea reversible y regulada.
Las quinasas añaden grupos fosfato; las fosfatasas los eliminan. La respuesta celular depende del equilibrio entre ambas actividades.
7.6. Vía fosfolipasa C — IP3/DAG — Ca²⁺/PKC
La vía de la fosfolipasa C suele activarse mediante receptores acoplados a proteína Gq.
La fosfolipasa C actúa sobre un fosfolípido de membrana llamado PIP2 o fosfatidilinositol 4,5-bisfosfato. Al hidrolizarlo, genera dos segundos mensajeros: IP3 y DAG.
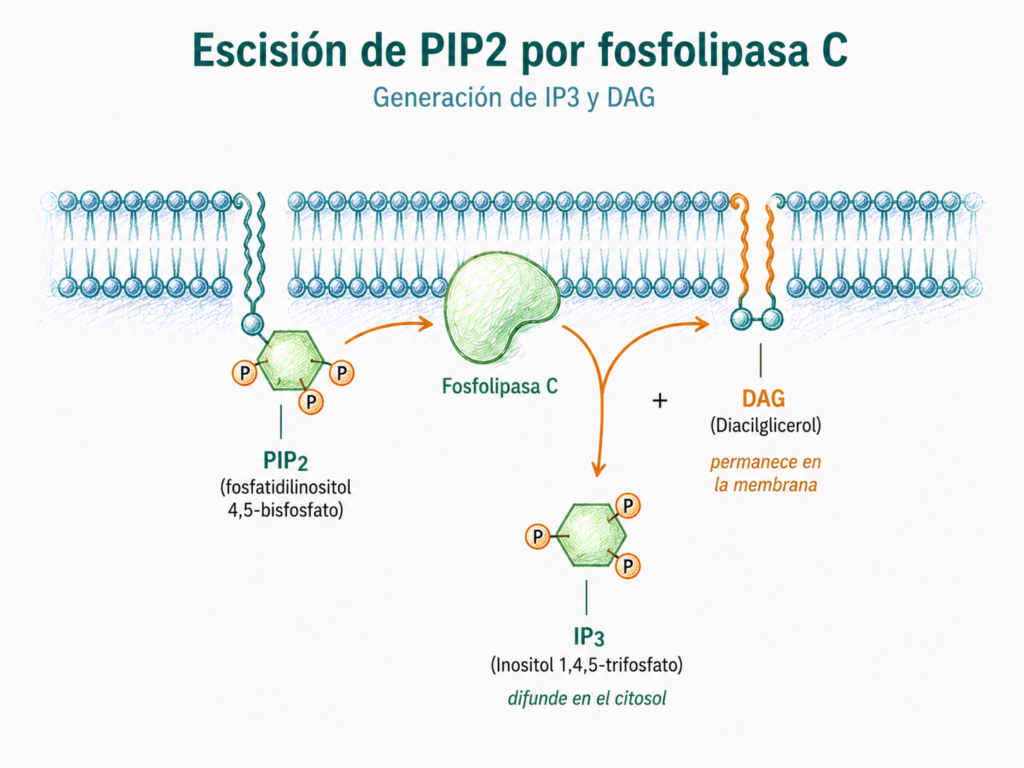
El IP3 es soluble y se desplaza por el citosol hasta el retículo endoplásmico. Allí se une a receptores específicos y favorece la salida de Ca²⁺ hacia el citoplasma.
El DAG permanece en la membrana plasmática. Junto con el Ca²⁺, contribuye a activar la proteína quinasa C o PKC.
| Segundo mensajero | Localización principal | Diana | Efecto |
|---|---|---|---|
| IP3 | Citosol | Receptor de IP3 en retículo endoplásmico | Libera Ca²⁺ al citosol |
| DAG | Membrana plasmática | Proteína quinasa C | Contribuye a activar PKC |
| Ca²⁺ | Citosol | Calmodulina · PKC · otras proteínas | Regula secreción, contracción, metabolismo y excitabilidad |
7.6.1. IP3, DAG y proteína quinasa C
El IP3 y el DAG actúan de forma coordinada, pero en lugares diferentes de la célula.
El IP3 moviliza Ca²⁺ desde el retículo endoplásmico. El aumento de Ca²⁺ citosólico activa proteínas sensibles al calcio, entre ellas la calmodulina y algunas quinasas.
El DAG permanece en la membrana y participa en la activación de PKC. Muchas formas de PKC requieren tanto DAG como Ca²⁺ para activarse.
La PKC fosforila proteínas diana en residuos de serina y treonina. Sus efectos son amplios y dependen del tipo celular. Puede participar en secreción, contracción, proliferación, respuesta inmune y regulación de la expresión génica.
La fosfolipasa C genera dos señales complementarias: IP3 moviliza Ca²⁺ y DAG ayuda a activar PKC.
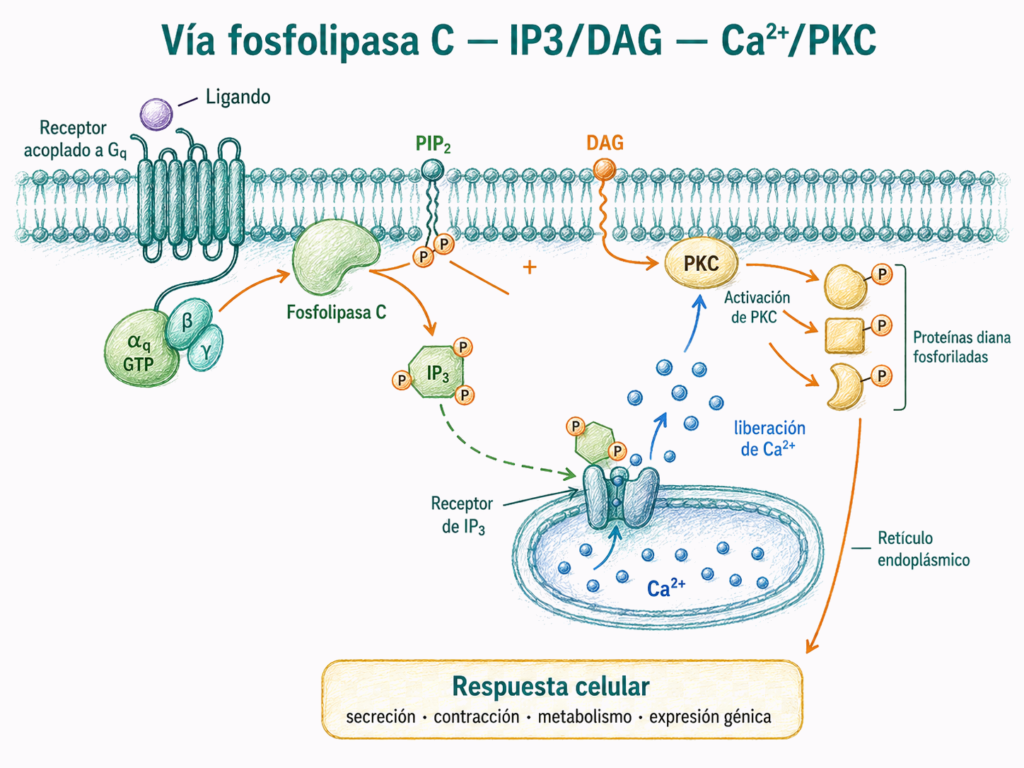
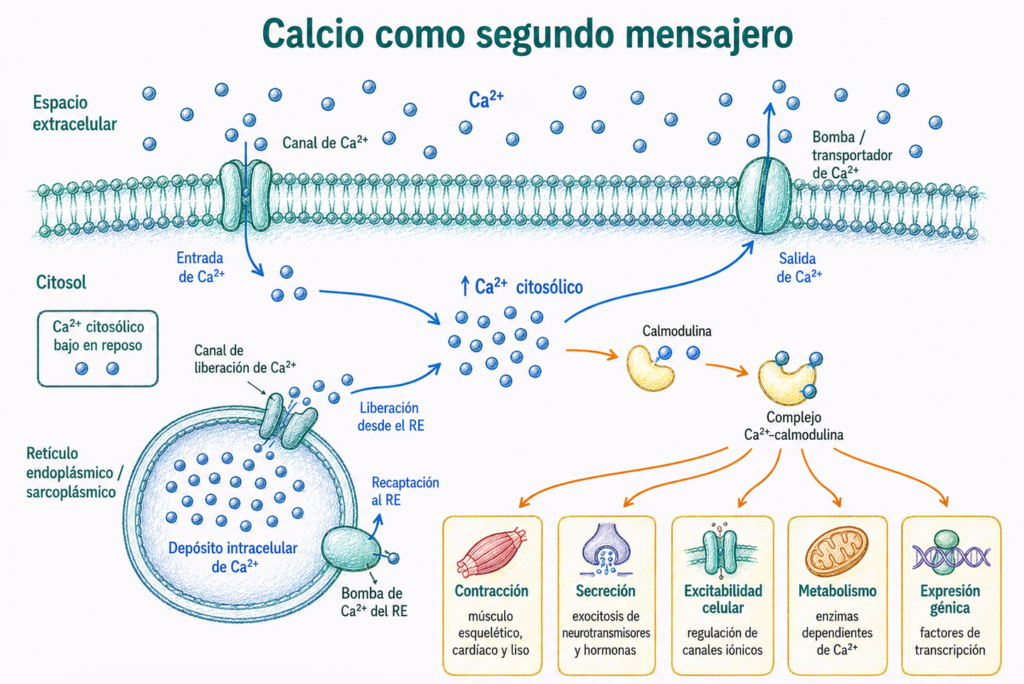
7.7. Calcio como segundo mensajero
El Ca²⁺ es uno de los segundos mensajeros más importantes de la célula.
A diferencia de otros mensajeros, no se sintetiza ni se degrada. La célula regula su señal modificando su entrada, su salida y su almacenamiento en compartimentos intracelulares.
En reposo, la concentración de Ca²⁺ libre en el citosol es muy baja. El Ca²⁺ se encuentra en mayor concentración en el exterior celular y en depósitos internos como el retículo endoplásmico o el retículo sarcoplásmico.
Cuando aumenta el Ca²⁺ citosólico, puede unirse a proteínas sensoras como la calmodulina. El complejo Ca²⁺-calmodulina regula enzimas, canales iónicos y proteínas relacionadas con contracción y secreción.
| Función regulada por Ca²⁺ | Ejemplo |
|---|---|
| Contracción | Músculo esquelético, cardíaco y liso |
| Secreción | Exocitosis de neurotransmisores y hormonas |
| Excitabilidad celular | Regulación de canales iónicos |
| Metabolismo | Activación o inhibición de enzimas dependientes de Ca²⁺ |
| Expresión génica | Activación de vías que regulan factores de transcripción |
El Ca²⁺ debe retirarse del citosol mediante bombas y transportadores. Si permanece elevado de forma excesiva, puede activar enzimas lesivas y contribuir al daño celular.
7.8. GMPc y óxido nítrico
El GMPc es otro segundo mensajero importante. Se forma a partir de GTP por acción de la guanilato ciclasa.
Una vía fisiológica relevante es la del óxido nítrico o NO. El NO es una molécula pequeña, liposoluble y de vida media corta. Difunde con facilidad hacia células vecinas y activa la guanilato ciclasa soluble.
El aumento de GMPc puede activar proteínas quinasas dependientes de GMPc y modificar canales o fosfodiesterasas. En el músculo liso vascular, esta vía favorece la relajación y contribuye a la vasodilatación.
La señal termina por degradación del GMPc mediante fosfodiesterasas específicas.
7.9. Regulación de la señal
7.9.1. Amplificación de la señal
Una característica fundamental de la señalización intracelular es la amplificación.
Una sola molécula de ligando puede activar un receptor durante un tiempo suficiente para estimular varias proteínas G. Cada proteína G puede activar un efector, y cada efector puede generar muchas moléculas de segundo mensajero.
Esto permite que señales extracelulares presentes en concentraciones muy bajas produzcan respuestas celulares amplias.
La amplificación es útil, pero exige mecanismos de control. Si la célula no pudiera apagar la señal, la respuesta sería excesiva o persistente.
7.9.2. Apagado e integración de la señal
La señalización intracelular debe ser reversible. La célula necesita responder, pero también necesita dejar de responder cuando el estímulo desaparece.
La señal puede apagarse por varios mecanismos:
- La subunidad α de la proteína G hidroliza GTP a GDP y se inactiva.
- Las fosfodiesterasas degradan AMPc o GMPc.
- Las bombas de Ca²⁺ retiran calcio del citosol.
- Las fosfatasas eliminan grupos fosfato de proteínas previamente fosforiladas.
- El receptor puede desensibilizarse si el estímulo persiste.
Además, las vías de señalización no actúan de forma aislada. Una célula puede recibir varias señales al mismo tiempo y combinarlas para generar una respuesta final. Esta integración depende del tipo celular, del receptor expresado, de los segundos mensajeros activados y del estado previo de la célula.
La célula no responde solo al mediador, sino al conjunto de señales que recibe y al contexto en el que las recibe.
7.10. Visión global de las principales vías
| Vía | Proteína G habitual | Efector | Segundo mensajero | Respuesta general |
|---|---|---|---|---|
| AMPc | Gs | Adenilato ciclasa | ↑ AMPc | Activación de PKA y fosforilación de proteínas |
| Inhibición de AMPc | Gi | Adenilato ciclasa | ↓ AMPc | Disminución de respuestas dependientes de PKA |
| IP3/DAG | Gq | Fosfolipasa C | IP3 · DAG · Ca²⁺ | Activación de PKC y vías dependientes de Ca²⁺ |
| GMPc | Variable | Guanilato ciclasa | GMPc | Relajación de músculo liso y regulación vascular |
