16.1. La sinapsis química: visión general
Las sinapsis químicas son el mecanismo predominante de comunicación neuronal en el ser humano. A diferencia de las sinapsis eléctricas — donde la señal se transmite directamente por corrientes iónicas — en la sinapsis química la señal eléctrica presináptica se convierte en una señal molecular: el neurotransmisor.
El principio fundamental es la conversión eléctrico-química-eléctrica: el potencial de acción presináptico desencadena la liberación de neurotransmisor, que difunde a través de la hendidura sináptica, se une a receptores específicos en la membrana postsináptica y genera una nueva respuesta eléctrica o metabólica en la célula postsináptica.
Los fundamentos de la sinapsis (concepto, tipos y organización morfofuncional) se desarrollaron en el tema 15. Los neurotransmisores específicos y sus receptores se desarrollan a partir del tema 17.
16.1.1. Tipos de neurotransmisores
Se han descrito más de 100 neurotransmisores distintos en el sistema nervioso. La clasificación más utilizada en fisiología los agrupa por peso molecular, lo que refleja diferencias en su mecanismo de síntesis, almacenamiento y cinética de acción.
Vía neuroquímica: secuencia de conexiones neuronales que utilizan un mismo neurotransmisor. Hablar de "vía colinérgica" o "vía dopaminérgica" implica que todas las neuronas de esa cadena comparten el mismo mensajero químico.
| Grupo | Familia | Ejemplos |
|---|---|---|
| Bajo peso molecular | Colinas | Acetilcolina |
| Catecolaminas | Adrenalina, noradrenalina, dopamina | |
| Otras aminas biógenas | Serotonina, histamina | |
| Aminoácidos | Glutamato, GABA, glicina | |
| Purinas | ATP, adenosina | |
| Alto peso molecular | Neuropéptidos | Encefalinas, sustancia P, VIP, oxitocina |
| No canónicos | Mensajeros lipídicos y gaseosos | NO, CO, endocannabinoides, ácido araquidónico |
Los neurotransmisores de bajo peso molecular se sintetizan directamente en el terminal presináptico a partir de precursores simples. Su síntesis es rápida y su disponibilidad se repone con facilidad durante la actividad sináptica intensa.
Los neuropéptidos se sintetizan en el soma neuronal como prepropéptidos y se procesan durante el transporte axonal hasta el terminal. Su síntesis es más lenta y costosa, lo que los hace más susceptibles al agotamiento ante estimulaciones prolongadas. Pueden actuar también como hormonas o neuromoduladores sistémicos.
Los no canónicos no siguen las reglas de la transmisión vesicular clásica: se sintetizan en la membrana o el citoplasma bajo demanda, difunden libremente a través de las membranas lipídicas y actúan frecuentemente de forma retrógrada. Se desarrollan en el T23 de esta asignatura.
16.2. Sinapsis canónicas y no canónicas
Las sinapsis químicas se dividen en dos grandes categorías según la dirección de la señal y el mecanismo de transmisión.
Las sinapsis canónicas siguen el patrón convencional: el neurotransmisor se libera desde la presinapsis, difunde a través de la hendidura y actúa sobre receptores en la postsinapsis. Son unidireccionales y utilizan receptores específicos. Constituyen la gran mayoría de las sinapsis del sistema nervioso.
Las sinapsis no canónicas invierten o alteran este esquema. El ejemplo más relevante son las sinapsis retrógradas, donde la señal viaja en sentido inverso (desde la postsinapsis hacia la presinapsis). Los mensajeros no canónicos más importantes son moléculas gaseosas o lipídicas que, por su naturaleza liposoluble, atraviesan libremente las membranas sin necesidad de receptores de superficie ni vesículas:
- Óxido nítrico (NO) y monóxido de carbono (CO): gases retrogradamente activos implicados en procesos de plasticidad sináptica y memoria.
- Ácido araquidónico y endocannabinoides: lípidos liposolubles que actúan retrogradamente sobre receptores presinápticos, modulando la liberación de neurotransmisor. El ácido araquidónico participa en procesos relacionados con el dolor y la inflamación.
Las sinapsis no canónicas no utilizan receptores de membrana en la célula origen del mensaje. El mensajero atraviesa directamente la membrana lipídica por difusión. Esto las hace fundamentalmente distintas de las sinapsis canónicas en términos de mecanismo, velocidad y regulación.
16.3. Ciclo del neurotransmisor
| Etapa | Localización | Mecanismo | Relevancia farmacológica |
|---|---|---|---|
| Síntesis | Terminal presináptico (o soma + transporte axonal) | Enzimas específicas a partir de precursores | Diana de fármacos que inhiben la síntesis (ej. inhibidores de DOPA descarboxilasa) |
| Almacenamiento | Vesículas sinápticas | Transportadores vesiculares concentran el NT en vesículas | Reserpina bloquea el almacenamiento de monoaminas |
| Liberación | Membrana presináptica | Exocitosis Ca²⁺-dependiente · liberación cuántica | Toxina botulínica bloquea la exocitosis; 4-aminopiridina la facilita |
| Unión al receptor | Membrana postsináptica | Unión transitoria y específica NT-receptor | Agonistas y antagonistas compiten por el receptor |
| Recaptación | Terminal presináptico y glía | Transportadores específicos de membrana | ISRS (fluoxetina), inhibidores de recaptación de NA (reboxetina) |
| Degradación enzimática | Hendidura sináptica / interior presináptico | Enzimas específicas (AChE, MAO, COMT) | Inhibidores de AChE (neostigmina), inhibidores de MAO (moclobemida) |
16.3.1. Síntesis y almacenamiento
La síntesis del neurotransmisor se produce en la neurona presináptica, generalmente en el propio terminal presináptico, aunque en algunos casos ocurre en el soma y el producto se transporta axonalmente hasta el terminal.
Una vez sintetizado, el neurotransmisor se almacena en vesículas sinápticas (orgánulos membranosos de ~40-50 nm de diámetro que lo protegen de la degradación enzimática citoplasmática y lo concentran para su liberación coordinada). En algunos casos lo que se almacena no es el neurotransmisor final sino sus precursores, que se procesan dentro de la vesícula o en el momento de la liberación.
16.3.2. Liberación en cuantos
La liberación del neurotransmisor ocurre en cuantos: paquetes discretos equivalentes al contenido de una vesícula sináptica. Cada cuanto representa la liberación simultánea de miles de moléculas de neurotransmisor por fusión de una vesícula con la membrana presináptica.
Unidad mínima de liberación de neurotransmisor, equivalente al contenido de una vesícula sináptica. La liberación cuántica garantiza que cada evento de exocitosis produce una cantidad reproducible y discreta de neurotransmisor en la hendidura sináptica.
Acoplamiento excitación-liberación: el papel del Ca²⁺
Cuando el potencial de acción llega al terminal presináptico, despolariza la membrana y abre los canales de calcio voltaje-dependientes presentes en la zona activa de la membrana presináptica. El Ca²⁺ entra a favor de gradiente electroquímico, sin gasto energético, y su aumento en el interior del terminal es la señal que desencadena la exocitosis de las vesículas.
El Ca²⁺ no empieza a entrar hasta la fase de repolarización del potencial de acción. Cuanto más tiempo dure el PA, más tiempo permanecen abiertos los canales y más Ca²⁺ entra, lo que se traduce en mayor liberación de vesículas. La energía que consume la neurona en este proceso se destina principalmente a la repolarización de la membrana mediante la bomba Na⁺/K⁺ ATPasa.
Los canales de calcio voltaje-dependientes se clasifican según su umbral de activación:
Bajo umbral (tipo T): se activan a voltajes cercanos al potencial de reposo (~−70 mV). Participan en la generación de ritmos oscilatorios.
Alto umbral: requieren mayor despolarización para abrirse. Se subdividen en:
- Tipo L: inactivación lenta. Presentes en músculo cardíaco y liso.
- Tipos N, P y Q: inactivación rápida. Son los más frecuentes en los terminales presinápticos del SNC.
Mecanismo molecular: proteínas SNARE
Las vesículas sinápticas se fusionan mediante un proceso mediado por un complejo proteico altamente específico denominado complejo SNARE.
Las vesículas almacenadas en el terminal están ancladas al citoesqueleto de actina mediante la proteína sinapsina. Cuando el Ca²⁺ entra en el terminal, activa la miosín kinasa calmodulina-dependiente a través del complejo Ca²⁺-calmodulina. Esta quinasa fosforila la sinapsina, que se disocia de la actina, liberando las vesículas hacia la membrana presináptica.
Una vez en la membrana, las vesículas se anclan y fusionan gracias al complejo SNARE, formado por:
- Sinaptobrevina (VAMP): proteína de la membrana de la vesícula
- Sintaxina y SNAP-25: proteínas de la membrana presináptica
- Sinaptotagmina: sensor de Ca²⁺ que activa la fusión cuando detecta el aumento de calcio
- α-SNAP, γ-SNAP y NSF: proteínas citosólicas reguladoras
La fusión genera un poro de fusión (similar en concepto a las uniones GAP) a través del cual el contenido de la vesícula se vierte a la hendidura sináptica. El proceso se cierra y la membrana vesicular queda integrada en la membrana presináptica.
La toxina botulínica (Botox) impide la formación del complejo SNARE bloqueando la exocitosis de acetilcolina en la unión neuromuscular y produciendo parálisis flácida. Se usa terapéuticamente en distonías, espasticidad y cosmética.
La toxina tetánica actúa de forma diferente: bloquea la liberación de GABA y glicina en las sinapsis inhibidoras de la médula espinal, produciendo parálisis espástica (tetania) por desinhibición de las neuronas motoras.
Reciclado de vesículas
Tras la exocitosis, la membrana vesicular fusionada con la membrana presináptica es recuperada por endocitosis y reciclada. La proteína clatrina forma una malla estructural (triskelia) que estabiliza las vesículas recuperadas y las dirige de vuelta al citoesqueleto de actina. La formación de vesículas estabilizadas con clatrina ocurre en ausencia de Ca²⁺ (cuando el calcio desciende tras el PA).
16.3.3. Unión al receptor
Tras su liberación, el neurotransmisor difunde a través de la hendidura sináptica y se une específicamente a receptores proteicos de la membrana postsináptica. La unión es específica. Cada neurotransmisor reconoce receptores complementarios en forma, carga y polaridad.
La unión neurotransmisor-receptor es transitoria: la constante de disociación garantiza que el neurotransmisor se disocie del receptor en un tiempo corto, permitiendo que la señal tenga una duración controlada y que el receptor quede disponible para nuevas señalizaciones. Una unión indefinida al receptor produciría una estimulación persistente. Esto es la base de algunas patologías y del mecanismo de acción de ciertos tóxicos.
16.3.4. Recaptación y degradación
Una vez disociado del receptor, el neurotransmisor debe ser eliminado de la hendidura sináptica para terminar la señal. Existen dos mecanismos principales:
- La recaptación. Es el transporte activo del neurotransmisor de vuelta a la neurona presináptica (o en algunos casos a la glía) mediante transportadores específicos de membrana. El NT recuperado puede reutilizarse, reempaquetándose en nuevas vesículas. La recaptación es el mecanismo de eliminación predominante para la mayoría de neurotransmisores y es la diana farmacológica de una gran familia de fármacos psiquiátricos, como los inhibidores de la recaptación de serotonina (ISRS), inhibidores de recaptación de noradrenalina, etc.
- La degradación enzimática destruye el neurotransmisor en la hendidura sináptica o en el interior de la neurona tras la recaptación. La enzima más conocida es la acetilcolinesterasa, que hidroliza la acetilcolina en la unión neuromuscular y en las sinapsis colinérgicas del SNC.
Las vesículas sinápticas también pueden ser recicladas tras la exocitosis: la membrana de la vesícula fusionada con la membrana presináptica es recuperada por endocitosis y reutilizada para formar nuevas vesículas.
La recaptación es un mecanismo de economía sináptica: el neurotransmisor recuperado se reutiliza, reduciendo el coste metabólico de la síntesis continua.
16.4. Tipos de sinapsis químicas
16.4.1. Según la velocidad: ionotrópicas vs. metabotrópicas
En la postsinapsis normalmente no se genera un potencial de acción directamente — lo que se produce es un cambio graduado del potencial de membrana (potencial postsináptico) que puede ser excitador o inhibidor. La excepción es la unión neuromuscular, donde la liberación masiva de acetilcolina genera directamente un PA en la célula muscular — se denomina sinapsis de activación directa.
Las sinapsis canónicas se clasifican según el tipo de receptor postsináptico activado, lo que determina la velocidad y el mecanismo de la respuesta:
- Las sinapsis ionotrópicas (o rápidas) activan receptores que son directamente canales iónicos. El canal y el receptor son la misma proteína. La unión del neurotransmisor abre el canal y permite el flujo inmediato de iones a través de la membrana postsináptica. La respuesta es muy rápida (inferior a 1 segundo). Son la base de la transmisión sináptica rápida en el SNC y en la unión neuromuscular.
- Las sinapsis metabotrópicas (o lentas) activan receptores acoplados a proteínas G. Las proteínas G son proteínas distintas del canal que, al unirse el NT, desencadenan cascadas de señalización intracelular mediadas por segundos mensajeros (AMPc, GMPc, IP₃, DAG, Ca²⁺) que modulan canales iónicos u otras proteínas efectoras de forma indirecta. La respuesta es más lenta (superior a 1 segundo), pero más duradera y con mayor capacidad de amplificación y modulación. Son especialmente relevantes en procesos de memoria y plasticidad sináptica a largo plazo.
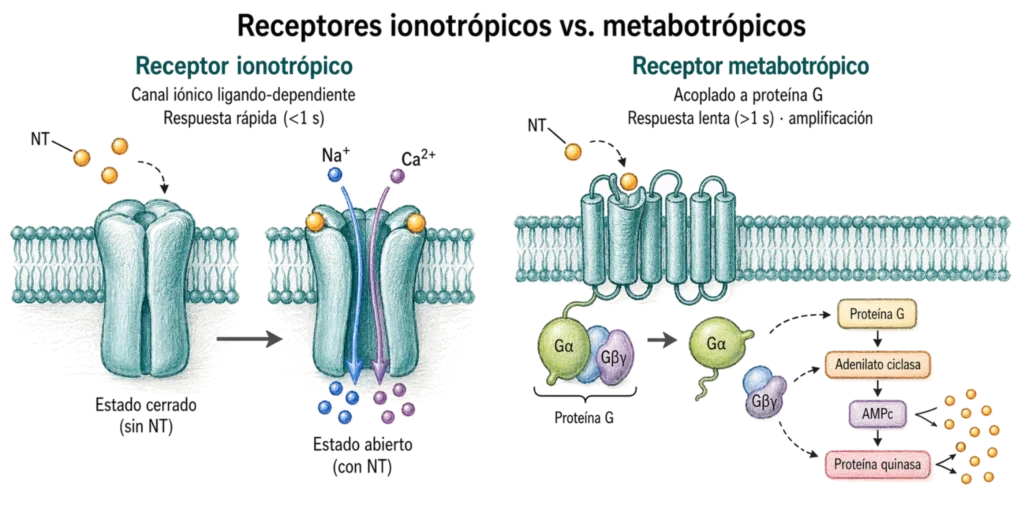
| Característica | Ionotrópica (rápida) | Metabotrópica (lenta) |
|---|---|---|
| Tipo de receptor | Canal iónico ligando-dependiente | Receptor acoplado a proteína G (GPCR) |
| Velocidad de respuesta | Rápida (< 1 segundo) | Lenta (> 1 segundo) |
| Mecanismo | Flujo iónico directo al abrirse el canal | Cascada de segundos mensajeros (AMPc, IP₃, DAG) |
| Duración del efecto | Breve | Prolongada |
| Amplificación de señal | No | Sí — un receptor activa múltiples proteínas G |
| Relevancia funcional | Transmisión rápida, reflejos, unión neuromuscular | Modulación, plasticidad, memoria |
| Ejemplos | nAChR (nicotínico), GABA-A, AMPA, NMDA | Receptores adrenérgicos β, mAChR (muscarínico), GABA-B |
16.4.2. Según la direccionalidad: dirigidas vs. no dirigidas
Las sinapsis canónicas también se clasifican según la precisión espacial de la transmisión:
- Las sinapsis dirigidas (o puntuales) establecen un contacto preciso entre terminal presináptico y membrana postsináptica con una hendidura sináptica reducida. El neurotransmisor actúa sobre un área pequeña y definida. Son las más frecuentes en el SNC y en el SNP somático. Se subdividen en sinapsis centrales (SNC) y periféricas (SNP).
- Las sinapsis no dirigidas tienen una separación mayor entre presinapsis y postsinapsis, superior a los 30 nm habituales. El neurotransmisor se libera de forma más difusa y puede actuar sobre receptores en un área más amplia. Son características del sistema nervioso autónomo (simpático y parasimpático), donde el neurotransmisor se libera desde varicosidades axónicas y difunde hacia células efectoras sin una hendidura sináptica especializada.
16.4.3. Cotransmisión
Durante décadas se asumió que cada neurona producía y liberaba un único tipo de neurotransmisor (la llamada ley de Dale). Esta afirmación ha sido refutada por la evidencia experimental: la mayoría de neuronas sintetizan y pueden liberar dos o más neurotransmisores simultáneamente desde la misma terminal presináptica.
Este fenómeno se denomina cotransmisión. Los neurotransmisores coexistentes pueden almacenarse en el mismo tipo de vesícula o en vesículas distintas, liberarse en proporciones variables según la frecuencia de estimulación, y actuar sobre los mismos o distintos receptores postsinápticos.
La cotransmisión añade una capa adicional de complejidad y flexibilidad a la señalización sináptica, permitiendo que una misma neurona produzca efectos diferentes en función del patrón de activación.
La ley de Dale en su formulación original ("una neurona, un neurotransmisor") es incorrecta. La reformulación moderna: "una neurona libera el mismo conjunto de neurotransmisores desde todos sus terminales" es más precisa pero tampoco absoluta.
16.5. Modulación presináptica
La liberación de neurotransmisor puede modularse directamente sobre el terminal presináptico, de forma independiente a la frecuencia de los potenciales de acción que llegan. Este mecanismo permite un ajuste fino de la transmisión sináptica.
Una neurona moduladora puede actuar sobre el terminal presináptico de otra neurona:
- Inhibición presináptica: la neurona moduladora hiperpolariza la membrana del terminal presináptico, reduciendo la probabilidad de apertura de los canales de Ca²⁺ y disminuyendo la liberación de neurotransmisor.
- Facilitación presináptica: la neurona moduladora despolariza parcialmente el terminal presináptico, aumentando la probabilidad de apertura de los canales de Ca²⁺ y facilitando la liberación de neurotransmisor.
16.6. Plasticidad sináptica a corto plazo
16.6.1. Potenciación postetánica
Tras una estimulación intensa y prolongada del terminal presináptico (estimulación tetánica), seguida de un período de reposo, la siguiente estimulación individual libera más vesículas de las que se liberaban antes de la estimulación tetánica. Este fenómeno se denomina potenciación postetánica.
El mecanismo implica la acumulación residual de Ca²⁺ intracelular y la inducción de enzimas que aumentan la síntesis y disponibilidad de neurotransmisor, incrementando la eficacia de la sinapsis durante un período posterior al tetano.
16.6.2. Agotamiento sináptico
La estimulación repetida a alta frecuencia puede agotar las reservas de vesículas disponibles para la liberación inmediata, produciendo una reducción progresiva de la respuesta postsináptica denominada agotamiento sináptico. Es un mecanismo de limitación intrínseca de la transmisión que previene la sobreestimulación.
16.7. Potenciales postsinápticos
La unión del neurotransmisor a su receptor postsináptico produce un cambio en la permeabilidad iónica de la membrana postsináptica: abre o cierra canales iónicos específicos. Esto altera el potencial de membrana de la célula postsináptica. Estos cambios de potencial se denominan potenciales postsinápticos (PPS) y son graduados, locales y decrementales. A diferencia del potencial de acción, no son regenerativos ni se propagan activamente.
16.7.1. Potencial postsináptico excitador (PPSE)
El PPSE es una despolarización local de la membrana postsináptica que aumenta la probabilidad de que se genere un potencial de acción.
El mecanismo más frecuente es la apertura de canales de Na⁺: el sodio entra a favor de su gradiente electroquímico, despolarizando la membrana desde el potencial de reposo (ej. de −70 mV a −69 mV). Esta pequeña despolarización por sí sola raramente es suficiente para alcanzar el umbral, necesita la suma de múltiples PPSE.
Las sinapsis excitadoras generan PPSE. Son las más frecuentes en las dendritas y espinas dendríticas.
16.7.2. Potencial postsináptico inhibidor (PPSI)
El PPSI es una hiperpolarización (o estabilización en el potencial de reposo) de la membrana postsináptica que disminuye la probabilidad de que se genere un potencial de acción.
Existen dos mecanismos principales:
- Apertura de canales de K⁺: el potasio sale a favor de su gradiente, hiperpolarizando la membrana.
- Apertura de canales de Cl⁻: el cloro entra a favor de su gradiente electroquímico, hiperpolarizando la membrana. Este mecanismo es especialmente frecuente en las sinapsis axo-somáticas inhibidoras. El GABA y la glicina actúan principalmente por esta vía.
Lo que determina si una sinapsis es excitadora o inhibidora no es el neurotransmisor en sí, sino el receptor al que se une. El mismo neurotransmisor puede producir PPSE en una célula y PPSI en otra, dependiendo del tipo de receptor postsináptico. La noradrenalina, por ejemplo, puede excitar o inhibir distintos tipos de músculo liso. El GABA, en cambio, actúa casi siempre sobre receptores que abren canales de Cl⁻, generando PPSI de forma consistente.
16.8. Integración sináptica
Una neurona recibe simultáneamente miles de señales sinápticas (excitadoras e inhibidoras) desde múltiples neuronas presinápticas. La integración sináptica es el proceso por el que la neurona suma todas estas señales y decide si genera o no un potencial de acción.
16.8.1. El cono axónico como punto de integración
Los PPSE e PPSI generados en dendritas y soma se propagan de forma pasiva (decremental) hasta el cono axónico (segmento inicial del axón). El cono axónico es el punto de integración final porque contiene la mayor densidad de canales de Na⁺ y K⁺ voltaje-dependientes de toda la neurona. Es el lugar donde el umbral de disparo es más fácil de alcanzar.
Si el sumatorio de potenciales que llega al cono axónico iguala o supera el potencial crítico de disparo, se genera un potencial de acción que se propaga por el axón. Si no lo alcanza, no se genera PA.
La localización de las sinapsis importa: las sinapsis más próximas al cono axónico tienen mayor influencia sobre el disparo que las sinapsis distales en las dendritas, cuyos potenciales se atenúan más al propagarse pasivamente.
16.8.2. Sumación espacial
La sumación espacial consiste en la activación simultánea de múltiples sinapsis en distintos puntos de la neurona. Los PPSE generados en distintas localizaciones se suman en el cono axónico. Si el sumatorio supera el umbral, se dispara el PA.
De forma análoga, la activación simultánea de sinapsis inhibidoras en distintos puntos suma PPSI que alejan la membrana del umbral, dificultando el disparo.
16.8.3. Sumación temporal
La sumación temporal ocurre cuando la misma sinapsis se activa repetidamente en rápida sucesión. Si el intervalo entre dos PPSE consecutivos es menor que el tiempo de decaimiento del potencial, el segundo PPSE se suma sobre el primero antes de que este se haya disipado, por lo que la despolarización acumulada puede alcanzar el umbral.
La integración sináptica es el mecanismo fundamental del procesamiento neuronal: cada neurona es un pequeño procesador que pondera continuamente miles de entradas excitadoras e inhibidoras y emite una salida binaria (dispara o no dispara). La complejidad del sistema nervioso emerge de la interacción de millones de estas decisiones simultáneas.
