19.1. Serotonina: generalidades y distribución
La serotonina (5-hidroxitriptamina, 5-HT) es una amina biógena con funciones muy diversas en el organismo. Solo el 1–2% de la serotonina corporal actúa como neurotransmisor en el SNC. La mayoría desempeña funciones no sinápticas: regula el tono del músculo liso vascular y aproximadamente el 8% se almacena en plaquetas, que la liberan durante la hemostasia para inducir vasoconstricción local y limitar la hemorragia.
En el SNC, las neuronas serotoninérgicas son poco abundantes pero muy ramificadas, lo que les permite establecer un número elevado de sinapsis y ejercer una influencia reguladora amplia y difusa. Se concentran en los núcleos del rafe del tronco encefálico, desde donde proyectan a prácticamente todas las regiones del encéfalo.
Las neuronas serotoninérgicas tienen un patrón de descarga muy lento (1–2 potenciales de acción por segundo). Esto se debe a que el potencial de acción en estas neuronas depende de canales de Na⁺ y Ca²⁺, lo que alarga la repolarización y prolonga los períodos refractarios.
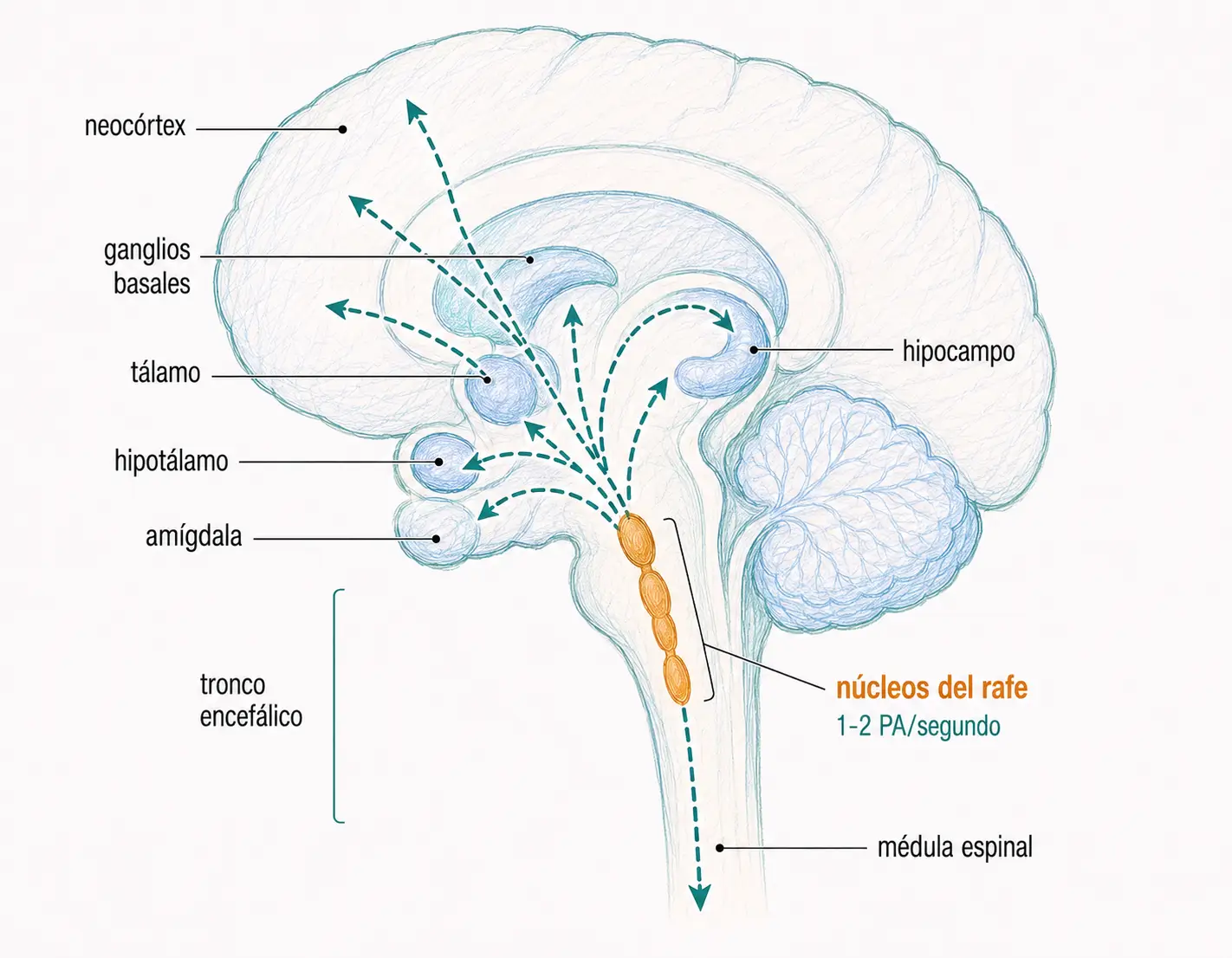
19.1.1. Funciones del sistema serotoninérgico
Las funciones de la serotonina en el SNC son variadas y afectan a múltiples dominios conductuales y vegetativos:
- Estado de ánimo y conducta: la serotonina participa en el mantenimiento del estado de ánimo equilibrado (eutimia). Su déficit se asocia a depresión; su alteración por drogas como el LSD (agonista parcial de receptores 5-HT2A) produce distorsiones perceptivas y alucinaciones.
- Atención y filtro sensorial: contribuye al filtrado de estímulos irrelevantes, permitiendo la focalización de la atención sobre una tarea concreta.
- Vigilia y tono encefálico: actúa como estimulador basal del SNC, manteniendo el estado de vigilia. Su descenso nocturno contribuye a la inducción del sueño.
- Funciones vegetativas: a través de proyecciones al hipotálamo, la serotonina influye en la regulación de la temperatura corporal, la ingesta de alimentos, la actividad sexual y la secreción hormonal (eje hipotálamo-hipofisario). Regula también la periodicidad de estas funciones; por ello, alteraciones del sueño pueden producir desregulación térmica y metabólica.
Los ISRS (inhibidores selectivos de la recaptación de serotonina: fluoxetina, sertralina, escitalopram) son el tratamiento de primera línea de la depresión y los trastornos de ansiedad. Al bloquear el transportador presináptico de serotonina, aumentan su concentración en la hendidura sináptica y prolongan su efecto postsináptico.
19.1.2. Síntesis y almacenamiento de serotonina
La serotonina se sintetiza a partir del aminoácido esencial triptófano (Trp), obtenido principalmente de la dieta. La ruta es de dos pasos:
| Paso | Sustrato → Producto | Enzima | Nota |
|---|---|---|---|
| 1 | Triptófano → 5-Hidroxitriptófano (5-HTP) | Triptófano hidroxilasa (TPH) | Paso limitante. Isoformas: TPH1 (periferia) y TPH2 (SNC). |
| 2 | 5-HTP → Serotonina (5-HT) | Aminoácido descarboxilasa aromática (AADC) | Misma enzima que convierte L-DOPA en dopamina (ver T18). |
Una vez sintetizada, la serotonina se empaqueta en vesículas sinápticas mediante el VMAT, el mismo transportador vesicular de monoaminas que almacena catecolaminas. Esto la protege de la degradación por MAO mitocondrial mientras permanece en el terminal.
La triptófano hidroxilasa tiene dos isoformas con distribución diferente: TPH1 se expresa en las células enterocromafines del intestino (fuente del 95% de la serotonina corporal) y en la glándula pineal; TPH2 se expresa exclusivamente en neuronas del SNC.
Esta distinción explica por qué los ISRS no afectan a la serotonina periférica de forma significativa.
19.1.3. Catabolismo de la serotonina
La serotonina libre en el citosol (no protegida en vesículas) es degradada por la MAO-A, anclada en la membrana externa mitocondrial. La vía es única:
Serotonina → ácido 5-hidroxiindolacético (5-HIAA) (enzima: MAO-A)
La degradación ocurre principalmente en las células gliales, donde la MAO-A tiene mayor expresión. En las propias neuronas serotoninérgicas, la MAO-B está presente pero tiene actividad muy baja sobre serotonina: la isoforma relevante para su catabolismo es siempre MAO-A.
El 5-HIAA se excreta por orina y su medición sirve como marcador indirecto de la actividad serotoninérgica. Está elevado en el síndrome carcinoide (tumor de células enterocromafines con hipersecreción de serotonina) y reducido en pacientes con depresión o tratados con IMAOs.
La MAO que degrada serotonina es MAO-A, no MAO-B. La MAO-B prefiere dopamina y β-feniletamina como sustratos. Esta distinción es farmacológicamente importante: la selegilina (inhibidor MAO-B selectivo) no bloquea la degradación de serotonina y por tanto tiene un perfil de interacciones muy diferente al de los IMAOs clásicos no selectivos.
19.1.4. Recaptación de serotonina
La terminación de la señal serotoninérgica depende principalmente de la recaptación por el transportador SERT (serotonin transporter, SLC6A4), presente en la membrana del terminal presináptico y en células gliales. SERT es dependiente de Na⁺ y Cl⁻, igual que NET y DAT para las catecolaminas.
La recaptación neuronal es autorregulatoria: la propia neurona presináptica recupera el neurotransmisor liberado, modulando la duración de la señal.
Los ISRS (fluoxetina, sertralina, escitalopram) bloquean selectivamente SERT. Los IMAOs producen un efecto complementario: al inhibir la MAO-A, impiden la degradación intracelular y aumentan el pool disponible para liberación. La combinación de ambos está contraindicada por riesgo de síndrome serotoninérgico.
El síndrome serotoninérgico es una emergencia médica por exceso de actividad 5-HT, típicamente por combinación de fármacos serotoninérgicos (ISRS + IMAO, ISRS + tramadol, ISRS + triptanos). Se presenta con la tríada: alteración del estado mental, inestabilidad autonómica e hiperreflexia/clonus.
El tratamiento incluye la retirada del fármaco causante y, en casos graves, ciproheptadina (antagonista 5-HT2A).
19.1.5. Melatonina: derivado serotoninérgico
La serotonina es precursora de la melatonina, hormona sintetizada en la glándula pineal. La pineal recibe señales luminosas desde la retina a través de una vía neuronal específica: cuando es de noche (ausencia de luz), convierte serotonina en melatonina mediante dos pasos enzimáticos adicionales (N-acetiltransferasa e hidroxiindol-O-metiltransferasa) y la libera a la circulación.
La melatonina regula el ritmo circadiano (ciclo vigilia/sueño) y sincroniza las funciones vegetativas con el ciclo luz/oscuridad. Dado que la serotonina es su precursor directo, alteraciones en la síntesis serotoninérgica repercuten sobre la producción de melatonina y, por tanto, sobre el sueño y los ritmos biológicos.
La regulación del ritmo circadiano por melatonina se desarrolla en el contexto de los sistemas generales de regulación (tema 30).
19.1.6. Receptores de serotonina
Existen siete familias de receptores de serotonina (5-HT1 a 5-HT7). Todos son metabotrópicos (GPCRs) salvo el 5-HT3, que es ionotrópico.
| Receptor | Tipo | 2.º mensajero / mecanismo | Relevancia |
|---|---|---|---|
| 5-HT1 (1A, 1B, 1D, 1E, 1F) | Metabotrópico | ↓ adenilato ciclasa → ↓ AMPc; apertura de canales de K⁺ | 5-HT1A: autorreceptor somatodendrítico. 5-HT1B/1D: diana de triptanos (migraña) |
| 5-HT2 (2A, 2B, 2C) | Metabotrópico | ↑ IP3/DAG (hidrólisis de fosfoinositol) | Diana del LSD (agonista 5-HT2A) y antipsicóticos atípicos (antagonistas 5-HT2A) |
| 5-HT3 | Ionotrópico | Canal catiónico (Na⁺/K⁺) → despolarización rápida | Único ionotrópico. Diana de antieméticos (ondansetrón) en quimioterapia |
| 5-HT4 | Metabotrópico | ↑ adenilato ciclasa → ↑ AMPc | Importante en motilidad gastrointestinal. Diana de procinéticos |
| 5-HT5 | Metabotrópico | ↓ adenilato ciclasa → ↓ AMPc | Expresión en hipocampo y corteza. Función en consolidación de memoria en estudio |
| 5-HT6 | Metabotrópico | ↑ adenilato ciclasa → ↑ AMPc | Modula glutamato y GABA en estriado y córtex. Implicado en cognición; diana activa en investigación para Alzheimer y esquizofrenia |
| 5-HT7 | Metabotrópico | ↑ adenilato ciclasa → ↑ AMPc | Regula ritmo circadiano y termorregulación. Diana de antipsicóticos atípicos (aripiprazol, lurasidona) |
El receptor 5-HT1A actúa como autorreceptor somatodendrítico: cuando detecta serotonina liberada por la propia neurona, hiperpolariza la membrana (↑ K⁺) y reduce su frecuencia de descarga. Es diana de los ansiolíticos como la buspirona y contribuye al mecanismo de adaptación a largo plazo de los ISRS.
El 5-HT3 es el único receptor ionotrópico de todo el sistema serotoninérgico. Su bloqueo por ondansetrón (antagonista 5-HT3) es la base del tratamiento antiemético en pacientes que reciben quimioterapia o radioterapia. Pregunta frecuente: "¿qué receptor de serotonina es ionotrópico?" → 5-HT3.
19.2. Histamina: generalidades, funciones y receptores
La histamina es una amina biógena cuyas funciones principales no son sinápticas. Actúa principalmente como autacoide: sustancia liberada localmente que actúa sobre células próximas a la secretora, sin necesidad de circulación sistémica.
Se sintetiza a partir del aminoácido histidina en un único paso enzimático:
Histidina → Histamina (enzima: histidina descarboxilasa, HDC)
La histamina sintetizada se almacena en gránulos citoplasmáticos de mastocitos y basófilos en la periferia, y en neuronas histaminérgicas del hipotálamo posterior (núcleo tuberomamilar) en el SNC.
A diferencia de las catecolaminas y la serotonina, la histamina no se almacena en vesículas sinápticas clásicas ni utiliza VMAT. En neuronas histaminérgicas se almacena en vesículas de núcleo denso y se libera por exocitosis Ca²⁺-dependiente, igual que otros neurotransmisores. En mastocitos, la liberación ocurre por desgranulación masiva ante estímulos alérgicos o inflamatorios.
19.2.1. Recaptación y degradación de histamina
La histamina no dispone de un transportador de recaptación neuronal eficiente, a diferencia de las catecolaminas (NET, DAT) o la serotonina (SERT). Su inactivación depende casi exclusivamente de la degradación enzimática, que sigue dos vías paralelas:
| Vía | Enzima | Localización | Producto |
|---|---|---|---|
| Metilación (vía principal en SNC) | Histamina N-metiltransferasa (HMT) | Citosol neuronal y glial | N-metilhistamina → degradada después por MAO-B |
| Oxidación (vía principal en periferia) | Diamino oxidasa (DAO) | Epitelio intestinal, placenta, riñón | Imidazolacetaldehído → ácido imidazolacético |
En el SNC predomina la vía HMT → MAO-B. En la periferia (intestino, placenta) predomina la DAO. La ausencia de recaptación eficiente hace que la histamina tenga una acción más difusa y prolongada que otros neurotransmisores, especialmente en tejidos con baja actividad enzimática.
La deficiencia o inhibición de DAO intestinal reduce la capacidad de degradar la histamina dietética (presente en pescado azul, queso curado, vino tinto, embutidos). La histamina acumulada se absorbe y produce síntomas sistémicos: rubor facial, cefalea, taquicardia, urticaria y, en casos graves, hipotensión.
Este cuadro se denomina intolerancia a la histamina y debe diferenciarse de la alergia alimentaria mediada por IgE.
19.2.2. Funciones de la histamina
Funciones periféricas de la histamina
Las funciones periféricas de la histamina son las más relevantes clínicamente:
- Permeabilidad vascular: aumenta la permeabilidad capilar, favoreciendo la extravasación de líquido y la formación de edema en los procesos inflamatorios y alérgicos.
- Vasodilatación: aumenta el calibre vascular produciendo hiperemia local.
- Músculo liso: regula la contracción del músculo liso bronquial (broncoconstricción en el asma) e intestinal.
- Secreción gástrica: estimula la secreción de ácido clorhídrico por las células parietales del estómago, actuando sobre receptores H2.
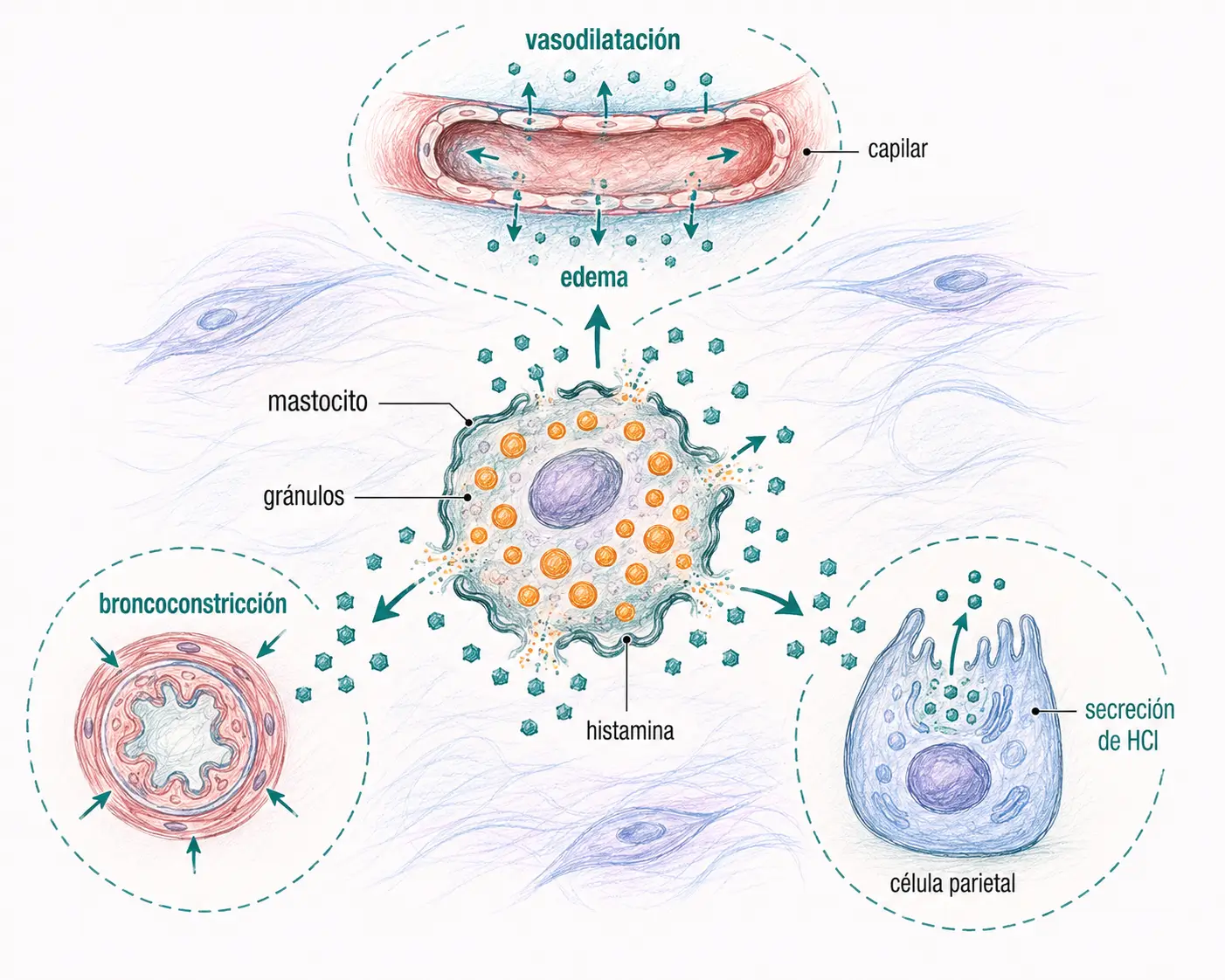
Funciones de la histamina en el SNC
Como neurotransmisor, la histamina regula múltiples funciones centrales a través de las neuronas del núcleo tuberomamilar del hipotálamo posterior, que proyectan difusamente a todo el encéfalo.
- Vigilia: las neuronas histaminérgicas del hipotálamo posterior promueven activamente el estado de vigilia. Su silenciamiento durante el sueño contribuye a la reducción del tono encefálico. Los antihistamínicos H1 de primera generación producen sedación por bloqueo de esta vía.
- Apetito y balance energético: la histamina inhibe la ingesta a través de receptores H1 en el núcleo ventromedial hipotalámico. Los antipsicóticos atípicos con antagonismo H1 central (olanzapina, quetiapina, clozapina) bloquean esta señal inhibidora y producen aumento de peso y síndrome metabólico, uno de sus efectos adversos más problemáticos a largo plazo.
- Señalización olfatoria y gustativa: la histamina participa en el procesamiento de estímulos químicos sensoriales en los bulbos olfatorios y en el tracto solitario.
Los antihistamínicos de primera generación (difenhidramina, clorfeniramina) cruzan la barrera hematoencefálica y bloquean receptores H1 centrales, produciendo sedación.
Los de segunda generación (loratadina, cetirizina) no cruzan la BHE y carecen de este efecto. Los antagonistas H2 (famotidina) reducen la secreción ácida gástrica en la úlcera péptica y el reflujo gastroesofágico. La ranitidina fue retirada del mercado en 2020 por contaminación con NDMA.
19.2.3. Receptores de histamina
Los receptores de histamina son cuatro, todos metabotrópicos (GPCRs):
| Receptor | Localización principal | 2.º mensajero | Función / relevancia clínica |
|---|---|---|---|
| H1 | Músculo liso, endotelio, SNC | ↑ IP3/DAG | Inflamación, alergia, broncoconstricción, vigilia. Diana de antihistamínicos de 1.ª y 2.ª generación |
| H2 | Células parietales gástricas, corazón | ↑ adenilato ciclasa → ↑ AMPc | Secreción ácida gástrica. Diana de antagonistas H2 (famotidina) |
| H3 | SNC (terminal presináptico) | ↓ adenilato ciclasa → ↓ AMPc | Autorreceptor presináptico: inhibe liberación de histamina y otros NT (GABA, ACh, DA). Diana del pitolisant (narcolepsia) |
| H4 | Células inmunes, médula ósea, intestino | ↓ adenilato ciclasa → ↓ AMPc | Quimiotaxis de eosinófilos y mastocitos. Implicado en prurito e inflamación crónica. Diana en investigación |
El siguiente diagrama recoge la clasificación completa de receptores del tema para facilitar la comparación entre ambos sistemas.
El receptor H3 es un autorreceptor presináptico heterólogo: no solo regula la liberación de histamina sino también la de otros neurotransmisores (GABA, acetilcolina, dopamina) en las terminales donde está presente. Su antagonismo aumenta la vigilia y la cognición, lo que explica el mecanismo del pitolisant en la narcolepsia.
Pregunta frecuente: "¿Cuál es el mecanismo de acción del pitolisant?" → Antagonista inverso del receptor H3.
Al bloquear el autorreceptor presináptico, aumenta la liberación de histamina endógena y promueve la vigilia. Primer fármaco no estimulante aprobado para narcolepsia (EMA 2016, FDA 2019).
