24.1. Organización morfofuncional del músculo esquelético
24.1.1. El sarcómero: unidad contráctil
La fibra muscular esquelética está recorrida longitudinalmente por miofibrillas, que son los elementos contráctiles de la célula. Al microscopio electrónico, las miofibrillas presentan una alternancia regular de bandas claras y oscuras que da al músculo esquelético su aspecto estriado característico.
- Las bandas A (anisotrópicas) son oscuras y corresponden a los filamentos gruesos de miosina, con los filamentos finos de actina parcialmente solapados en sus extremos.
- En el centro de la banda A se encuentra la zona H (o banda H), una región más clara donde los filamentos finos no llegan y solo existen filamentos gruesos.
- En el centro de la zona H se distingue la línea M, estructura de anclaje transversal de los filamentos gruesos entre sí, que mantiene su disposición hexagonal.
- Las bandas I (isotrópicas) son claras y contienen únicamente filamentos finos.
- En el centro de cada banda I se encuentra la línea Z, estructura de anclaje de los filamentos de actina.
- La titina (o conectina) es la tercera proteína filamentosa del sarcómero: une la línea M con la línea Z siguiendo el filamento grueso como un muelle molecular. Aporta elasticidad pasiva al sarcómero y evita el sobreestiramiento de la miofibrilla.
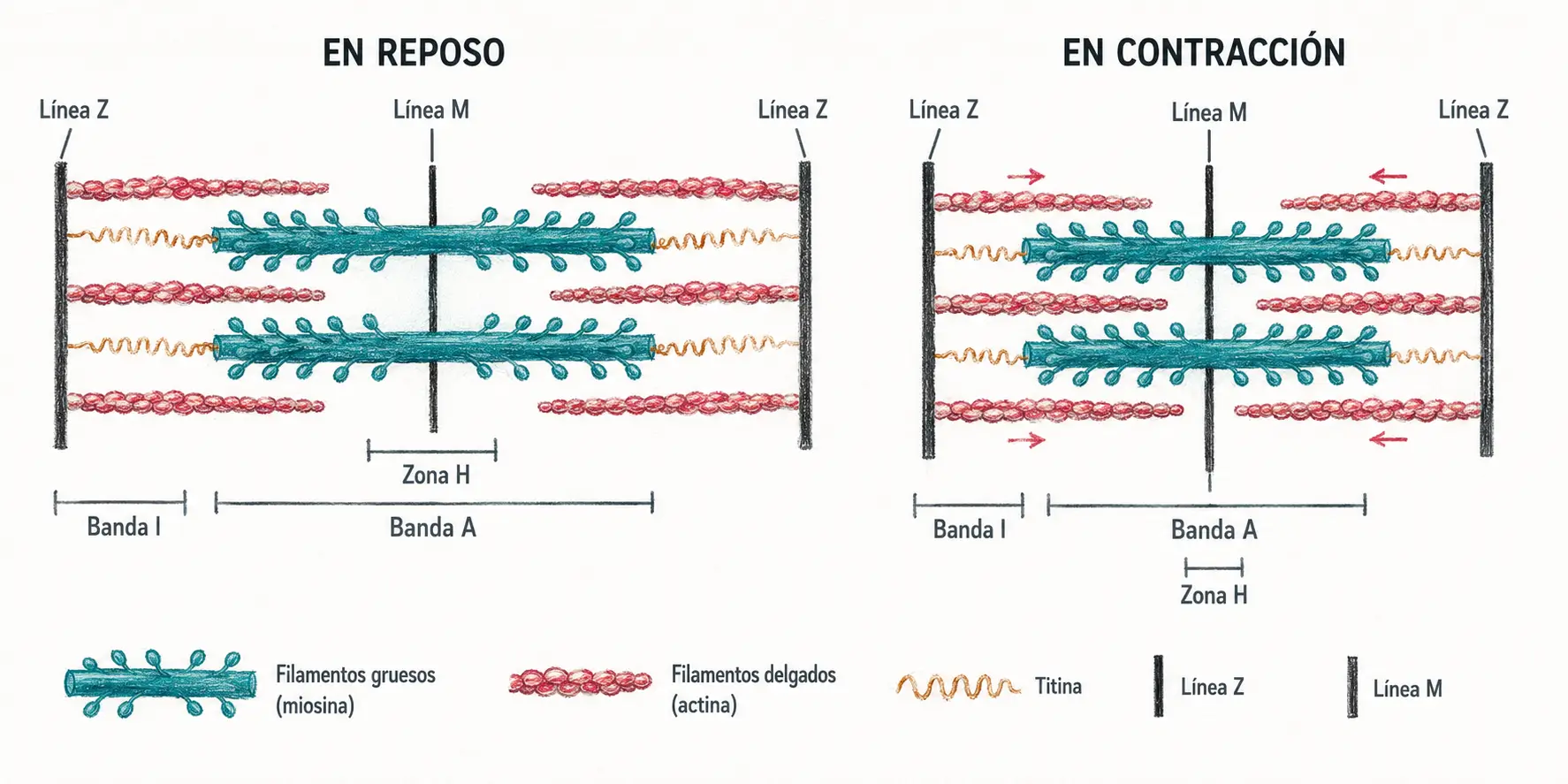
Sarcómero: unidad contráctil de la miofibrilla, delimitada por dos líneas Z consecutivas. Contiene en su centro la banda A con los filamentos gruesos, y en sus extremos las semibandas I con los filamentos finos.
Cuando el sarcómero se contrae, los filamentos finos se deslizan hacia el interior, la banda I se acorta, las líneas Z se aproximan y el sarcómero en conjunto acorta su longitud. Los filamentos en sí no cambian de longitud: es el grado de solapamiento lo que varía. Este modelo se conoce como teoría del filamento deslizante.
La ultraestructura detallada del sarcómero, la organización tridimensional de los filamentos y la morfología del retículo sarcoplásmico se estudian en Histología General (34443), Tema 12 — Tejido muscular I: músculo estriado esquelético.
24.1.2. Composición molecular de los filamentos
Los filamentos gruesos están formados por polímeros de miosina II. Cada monómero de miosina está formado por dos cadenas pesadas. La región globular constituye las cabezas (fragmento S1) y la región helicoidal enrollada forma la cola (fragmentos S2 y LMM). Asociadas al cuello de cada cabeza hay dos cadenas ligeras (una esencial y una reguladora), que modulan la actividad ATPasa pero no forman la cola. Las cabezas de miosina tienen dos propiedades esenciales:
- Actividad ATPasa (hidrolizan ATP para obtener energía).
- Capacidad de unirse a la actina.
La troponina es el interruptor molecular de la contracción: cuando no hay Ca²⁺, la tropomiosina bloquea los sitios de unión de la actina y el músculo permanece relajado. Cuando el Ca²⁺ se une a la troponina C, el interruptor se activa.
El conjunto formado por la cabeza y el cuello de la miosina constituye el puente cruzado, que es el elemento que genera la fuerza contráctil mediante un movimiento de vaivén sobre la cola.
Los filamentos finos están formados por tres proteínas:
- Actina F: polímero helicoidal formado por la polimerización de actina G (globular). Dos cadenas de actina F se enrollan entre sí formando la columna vertebral del filamento.
- Tropomiosina: proteína fibrilar que corre a lo largo del surco del doble helix de actina, cubriendo los sitios de unión para la miosina en estado de reposo.
- Troponina: complejo de tres subunidades con funciones diferenciadas:
- La troponina T ancla el complejo a la tropomiosina.
- La troponina C se une al Ca²⁺ con alta afinidad.
- La troponina I inhibe la ATPasa de la actomiosina y estabiliza la posición bloqueante de la tropomiosina sobre los sitios de unión de la actina en ausencia de Ca²⁺.
24.2. Fenómenos eléctricos en el músculo: el potencial de acción muscular
La fibra muscular esquelética es una célula excitable. Tiene un potencial de membrana en reposo de entre −80 y −90 mV, algo más negativo que el de la neurona típica. Cuando el potencial de placa motora supera el umbral, se dispara un potencial de acción que se propaga por todo el sarcolema.
24.2.1. Características del potencial de acción muscular
El potencial de acción muscular sigue las mismas fases básicas que el neuronal: despolarización rápida por apertura de canales de Na⁺ voltaje-dependientes, seguida de repolarización por apertura de canales de K⁺. Sin embargo, presenta una diferencia importante: la duración es mayor, de 1 a 5 ms frente al milisegundo típico del neuronal.
Esta mayor duración se debe a la cinética más lenta de los canales de K⁺ de repolarización en la membrana muscular. Los canales de Ca²⁺ tipo L (los mismos DHPR que actúan como sensores de voltaje en los túbulos T) contribuyen de forma menor a la corriente de entrada durante el potencial de acción, pero no generan una meseta prolongada ni aportan Ca²⁺ significativo para la contracción.
En el músculo esquelético, el Ca²⁺ que activa la maquinaria contráctil proviene casi exclusivamente del retículo sarcoplásmico, liberado a través de los receptores RyR1. El Ca²⁺ extracelular no es esencial para desencadenar la contracción esquelética, a diferencia de lo que ocurre en el músculo cardíaco.
La meseta prolongada del potencial de acción por entrada de Ca²⁺ extracelular es característica del músculo CARDÍACO, no del esquelético. En el músculo esquelético el Ca²⁺ contráctil proviene del retículo sarcoplásmico (RyR1). Confundir ambos mecanismos es uno de los errores más frecuentes en los exámenes de este bloque.
| Parámetro | PA neuronal | PA muscular esquelético | PA muscular cardíaco |
|---|---|---|---|
| Duración | ~1 ms | 1–5 ms | 200–400 ms |
| Potencial de reposo | −70 mV | −80 a −90 mV | −90 mV |
| Corriente de entrada principal | Na⁺ (Nav) | Na⁺ (Nav) | Na⁺ (Nav) + Ca²⁺ (Cav tipo L) |
| Meseta por Ca²⁺ extracelular | No | No | Sí (fase 2) |
| Ca²⁺ contráctil | — | Retículo sarcoplásmico (RyR1) | RS (RyR2) + Ca²⁺ extracelular (CICR) |
| Sensor de voltaje → Ca²⁺ | — | DHPR → RyR1 (mecánico directo) | DHPR → entrada Ca²⁺ → RyR2 (CICR) |
| Período refractario | Corto (~1 ms) | Corto (permite tétanos) | Largo (impide tétanos cardíaco) |
| Velocidad de conducción | 70–120 m/s (fibra A mielínica) | 3–5 m/s | 0,3–1 m/s (miocardio ventricular) |
La duración mayor del potencial de acción muscular no es un defecto: es funcionalmente coherente con la necesidad de desencadenar un proceso mecánico (la contracción) que requiere tiempo para activarse. El potencial neuronal solo necesita transmitir información; el muscular necesita además poner en marcha maquinaria contráctil.
24.2.2. Propagación por el sarcolema y los túbulos T
Una vez generado, el potencial de acción se propaga en ambos sentidos desde la placa motora hacia los extremos de la fibra. La velocidad de conducción es unas diez veces menor que en una fibra nerviosa mielínica, del orden de 3-5 m/s.
La propagación no se limita a la superficie del sarcolema. Los túbulos T son invaginaciones tubulares del sarcolema que penetran transversalmente en el interior de la fibra, llevando el potencial de acción hasta las proximidades del retículo sarcoplásmico en toda la sección transversal de la célula. Sin los túbulos T, el potencial de acción tardaría demasiado en alcanzar las miofibrillas centrales por difusión pasiva.
El mecanismo por el que el potencial de acción en el túbulo T desencadena la liberación de Ca²⁺ desde el retículo sarcoplásmico constituye el acoplamiento excitación-contracción, que se desarrolla en el apartado 3 de este tema.
En el punto donde el túbulo T queda flanqueado por dos cisternas terminales del retículo sarcoplásmico se forma la tríada: la unidad estructural donde el DHPR del túbulo T contacta físicamente con el RyR1 de las cisternas adyacentes. Es en la tríada donde el potencial eléctrico se convierte en señal mecánica que abre el canal de Ca²⁺.
La estructura del sarcolema, los túbulos T y el retículo sarcoplásmico se describe morfológicamente en Histología General (34443), Tema 12 — Tejido muscular I.
24.3. Proceso de la contracción muscular: acoplamiento excitación-contracción
El término acoplamiento excitación-contracción designa la secuencia de eventos que une el potencial de acción eléctrico con la respuesta mecánica de la miofibrilla. El eslabón central es el Ca²⁺.
24.3.1. Liberación de Ca²⁺ desde el retículo sarcoplásmico
El potencial de acción que se propaga por el sarcolema penetra en el interior de la fibra a través de los túbulos T (descritos en el apartado 2). En la membrana del túbulo T hay receptores de dihidropiridina (DHPR) sensores de voltaje que detectan el cambio de potencial.
Estos DHPR están físicamente acoplados a los receptores de rianodina (RyR1) de la membrana del retículo sarcoplásmico adyacente. Cuando el sensor de voltaje se activa, transmite el cambio conformacional al RyR1, que se abre y libera Ca²⁺ almacenado al sarcoplasma de manera masiva y sincronizada.
Este acoplamiento es de naturaleza mecánica directa: el cambio conformacional del DHPR se transmite físicamente al RyR1 sin necesidad de un segundo mensajero. Es lo que distingue al músculo esquelético del cardíaco, donde la apertura del RyR2 depende del Ca²⁺ extracelular que entra durante el potencial de acción (Ca²⁺-induced Ca²⁺ release o CICR).
En el músculo cardíaco, el Ca²⁺ que entra durante el potencial de acción actúa como disparador del RyR2 (CICR). Por eso el músculo cardíaco depende del Ca²⁺ extracelular. Si se suprime, la contracción cardíaca falla. En el músculo esquelético, la contracción puede ocurrir aunque no haya Ca²⁺ en el exterior, porque el DHPR actúa como sensor de voltaje puro.
El Ca²⁺ liberado empapa las miofibrillas circundantes, alcanzando concentraciones que pasan de ~100 nM en reposo a ~10 µM durante la activación.
24.3.2. Mecanismo molecular del golpe de remo
Con el Ca²⁺ presente en el sarcoplasma, la secuencia contráctil se desarrolla en cuatro pasos cíclicos:
- El Ca²⁺ se une a la troponina C, produciendo un cambio conformacional que arrastra a la troponina T y desplaza la tropomiosina fuera del surco, dejando accesibles los sitios de unión de la actina.
- La cabeza de miosina, que lleva unido ADP + Pi tras la hidrólisis previa de ATP, se une al sitio activo de la actina, formando el puente cruzado.
- La liberación del Pi desencadena el golpe de remo: la cabeza de miosina rota sobre su cuello, desplazando el filamento de actina hacia el centro del sarcómero. Al final del golpe se libera el ADP.
- Una nueva molécula de ATP se une a la cabeza de miosina, provocando su desprendimiento de la actina. La ATPasa hidroliza el ATP en ADP + Pi, recargando la cabeza para el siguiente ciclo.
El ATP tiene dos papeles en la contracción: aportar la energía del golpe de remo y permitir el desprendimiento de la cabeza de miosina de la actina. Sin ATP, los puentes cruzados quedan atrapados en estado unido: es la base molecular del rigor mortis.
Un error frecuente es pensar que el Ca²⁺ genera directamente el movimiento. No es así: el Ca²⁺ levanta el bloqueo de la tropomiosina, pero la fuerza la produce la hidrólisis de ATP en la cabeza de miosina. El Ca²⁺ es el interruptor; el ATP es el combustible.
Rigor mortis: tras la muerte, cesa la producción de ATP. Sin ATP, las cabezas de miosina no pueden desprenderse de la actina tras el golpe de remo: los puentes cruzados quedan atrapados en estado unido y el músculo se vuelve rígido. Esta rigidez se mantiene hasta que la degradación proteolítica progresiva de actina y miosina disuelve los puentes cruzados y la rigidez desaparece.
24.4. Relajación muscular
La relajación se inicia cuando cesa la llegada de potenciales de acción a la fibra muscular. Sin estimulación, los canales RyR1 se cierran y deja de liberarse Ca²⁺ al sarcoplasma.
El Ca²⁺ residual es reintroducido activamente en el retículo sarcoplásmico por la bomba SERCA (Ca²⁺-ATPasa del retículo sarco/endoplásmico), que hidroliza ATP para transportar dos iones de Ca²⁺ por molécula de ATP consumida, contra gradiente de concentración.
A medida que la concentración de Ca²⁺ en el sarcoplasma cae por debajo del umbral de activación de la troponina C, la tropomiosina vuelve a cubrir los sitios activos de la actina, los puentes cruzados no pueden formarse y el músculo se relaja.
La relajación muscular tiene coste energético. Tanto la contracción (golpe de remo) como la relajación (bombeo de Ca²⁺ al RS) consumen ATP. Un músculo en actividad continua agota ATP en ambas fases del ciclo.
24.5. Consumo de energía en la contracción y la relajación
El músculo en actividad tiene una demanda energética elevada y cambiante. Para cubrirla dispone de cuatro fuentes de ATP que se activan en cascada según la duración e intensidad del esfuerzo.
El ATP almacenado en la fibra muscular es la fuente directa e inmediata, pero su concentración es escasa (unos 5 µmol por gramo de músculo): suficiente solo para mantener una contracción máxima durante 1 o 2 segundos.
La fosfocreatina (PCr) es la reserva de alta energía más rápida. La enzima creatina quinasa transfiere el grupo fosfato de la fosfocreatina al ADP regenerando ATP en milisegundos, sin necesidad de oxígeno ni de activar vías metabólicas complejas. Esta reserva se agota en 10-15 segundos de esfuerzo máximo.
La glucólisis anaeróbica permite regenerar ATP a partir del glucógeno muscular o de la glucosa sanguínea a una velocidad elevada. Su producto final es el lactato, que se acumula en el músculo y en sangre. Es la fuente predominante en esfuerzos de alta intensidad que duran entre 10 segundos y 2 minutos.
La fosforilación oxidativa (cadena de transporte de electrones mitocondrial) es la fuente de mayor rendimiento energético: produce hasta 30-32 moléculas de ATP por mol de glucosa. Requiere oxígeno y tarda más en activarse, pero es insustituible en esfuerzos prolongados de intensidad moderada-baja.
Las cuatro fuentes de ATP se activan en cascada: ATP directo → fosfocreatina → glucólisis → fosforilación oxidativa. Cuanto mayor es la intensidad del esfuerzo, más peso tienen las fuentes anaeróbicas; cuanto mayor es la duración, más peso tiene la vía oxidativa.
El tipo de fibra muscular condiciona qué fuente predomina. Las fibras tipo I (lentas, oxidativas) dependen fundamentalmente de la fosforilación oxidativa y son resistentes a la fatiga. Las fibras tipo IIa (rápidas, oxidativo-glucolíticas) combinan ambas rutas. Las fibras tipo IIx (rápidas, glucolíticas) dependen de la glucólisis anaeróbica y se fatigan rápidamente. La correspondencia entre tipos de fibra y función mecánica se desarrolla en el Tema 25 · Mecánica y electrofisiología muscular.
La fatiga muscular es la disminución reversible de la capacidad de generar fuerza durante la actividad sostenida. Sus causas son múltiples: agotamiento de las reservas de fosfocreatina y glucógeno, acidosis intracelular por acumulación de H⁺ (asociada al lactato), acumulación de Pi inorgánico que interfiere con el ciclo de puentes cruzados y, en esfuerzos prolongados, alteraciones en la disponibilidad de Ca²⁺ del retículo sarcoplásmico.
La comprensión del metabolismo energético muscular tiene aplicación directa en patología: las glucogenosis musculares (como la enfermedad de McArdle, por déficit de glucógeno fosforilasa) impiden la glucólisis anaeróbica y producen calambres y mioglobinuria con el ejercicio. Las miopatías mitocondriales alteran la fosforilación oxidativa y cursan con intolerancia al ejercicio y debilidad progresiva.
| Fuente de ATP | Velocidad | Duración | Requiere O₂ | Producto final |
|---|---|---|---|---|
| ATP directo | Inmediata | 1–2 s | No | ADP + Pi |
| Fosfocreatina (PCr) | Muy rápida | 10–15 s | No | Creatina + ATP |
| Glucólisis anaeróbica | Rápida | 10 s – 2 min | No | Lactato + ATP |
| Fosforilación oxidativa | Lenta (activación) | Minutos – horas | Sí | CO₂ + H₂O + ATP (×30–32) |
