12.1. El nervio como cable funcional
El sistema nervioso necesita que la información viaje de forma rápida, fiable y sin mezclar señales de distinta naturaleza. Para ello, cada fibra nerviosa conduce un tipo específico de información por una vía dedicada: esto es el principio de la línea marcada. Lo que el cerebro interpreta no depende de la fibra que se activa, sino del destino al que llega esa señal. Si estimulas artificialmente una fibra que normalmente conduce tacto, percibirás tacto aunque el estímulo haya sido eléctrico.
Las fibras nerviosas no son todas iguales. Difieren en diámetro, en si están o no envueltas en mielina, y en la velocidad a la que propagan el potencial de acción. Estas tres características están correlacionadas y son la base de las clasificaciones que se usan en fisiología.
Fibra nerviosa: prolongación neuronal especializada en conducir potenciales de acción. Sus propiedades funcionales dependen del diámetro, la mielinización y la organización de sus canales iónicos.
La conducción del potencial de acción, incluidos los mecanismos de corrientes locales y conducción saltatoria que explican por qué la mielina aumenta la velocidad, se desarrollan en el T11 — Conducción del potencial de acción de esta misma asignatura.
12.2. Clasificación de las fibras nerviosas
12.2.1. Criterios de clasificación de las fibras nerviosas
Tres parámetros permiten clasificar cualquier fibra nerviosa:
- Presencia de vaina de mielina: mielínicas o amielínicas.
- Diámetro axonal: medido en micrómetros (μm). A mayor diámetro, menor resistencia al flujo de corriente longitudinal.
- Velocidad de conducción: medida en metros por segundo (m/s). Se correlaciona directamente con el diámetro y con la mielinización.
Los tres parámetros van de la mano: las fibras más gruesas tienden a ser mielínicas y las más rápidas. No son criterios independientes, sino facetas distintas de una misma realidad estructural.
12.2.1. Clasificación de Erlanger-Gasser (fibras A, B y C)
Esta es la clasificación general, aplicable a cualquier fibra nerviosa, sea motora, sensitiva o autonómica. La propusieron Joseph Erlanger y Herbert Gasser en los años 1920-1930 a partir de registros de velocidad de conducción en nervios compuestos, trabajo que les valió el Premio Nobel de Fisiología en 1944.
Las fibras se dividen en tres grupos principales: A, B y C.
- Las fibras A son fibras mielínicas de gran diámetro. Se subdividen en cuatro subtipos según diámetro y velocidad decrecientes: α, β, γ y δ.
- Las fibras Aα son las más gruesas (10-20 μm) y las más rápidas (70-120 m/s). Incluyen las motoneuronas alfa que inervan fibras musculares extrafusales y las aferentes primarias del huso muscular. Su velocidad de conducción elevada es imprescindible para los reflejos motores que requieren latencias muy cortas.
- Las fibras Aβ (8-14 μm, 35-80 m/s) transmiten información táctil de umbral bajo: tacto fino, presión, vibración. Son las que activan los mecanorreceptores cutáneos de adaptación rápida y lenta.
- Las fibras Aγ (5-9 μm, 10-50 m/s) inervan las fibras intrafusales del huso muscular, modulando su sensibilidad. No generan fuerza directamente, pero regulan la ganancia del reflejo miotático.
- Las fibras Aδ (<5 μm, <30 m/s) son las más delgadas dentro del grupo A. Transmiten dolor agudo y bien localizado (el primer dolor que se siente tras un pinchazo) y temperatura fría. Su fina capa de mielina les da velocidad suficiente para esta función de alerta rápida.
- Las fibras B (1-3 μm, 3-15 m/s) son fibras mielínicas exclusivas de las vías preganglionares del sistema nervioso autónomo.
- Las fibras C: (0,5-2 μm, 0,5-2 m/s) son fibras amielínicas. Son las más delgadas y las más lentas. Transmiten dolor sordo y difuso (el segundo dolor, de inicio tardío tras una lesión), temperatura cálida y señales autonómicas posganglionares.
| Fibra | Mielina | Diámetro (μm) | Velocidad (m/s) | Función principal |
|---|---|---|---|---|
| Aα | Sí | 10–20 | 70–120 | Motoneuronas α; aferentes del huso muscular (primarias) |
| Aβ | Sí | 8–14 | 35–80 | Tacto fino, presión, vibración |
| Aγ | Sí | 5–9 | 10–50 | Motoneuronas γ; regulación del huso muscular |
| Aδ | Sí (fina) | <5 | <30 | Dolor agudo localizado; temperatura fría |
| B | Sí (fina) | 1–3 | 3–15 | Autonómicas preganglionares |
| C | No | 0,5–2 | 0,5–2 | Dolor sordo difuso; temperatura cálida; autonómicas posganglionares |
A mayor diámetro axonal y mayor grado de mielinización, mayor velocidad de conducción. Las fibras Aα, con 70-120 m/s, son unas 100 veces más rápidas que las fibras C amielínicas. Esta diferencia tiene consecuencias funcionales directas: el "primer dolor" (agudo, localizado, inmediato) viaja por Aδ; el "segundo dolor" (sordo, difuso, tardío) viaja por C.
12.2.2. Clasificación de Lloyd-Hunt (grupos I–IV)
Esta clasificación, propuesta por Lloyd y Hunt en los años 1940, se aplica exclusivamente a las fibras aferentes sensitivas de los nervios musculares y cutáneos. Es la nomenclatura habitual en fisiología sensorial y en el estudio de los reflejos espinales.
Ambas nomenclaturas describen las mismas fibras desde perspectivas distintas. Erlanger-Gasser clasifica por velocidad de conducción y se aplica a cualquier fibra. Lloyd-Hunt clasifica por origen receptor y se aplica solo a aferentes sensitivas. En la práctica, los fisiólogos usan las dos y conviene saber relacionarlas.
- Las fibras del grupo I son las más gruesas y rápidas de las sensitivas. Se subdividen en dos:
- Grupo Ia (o Iα): 20 μm, ~120 m/s. Proceden de las terminaciones anuloespirales del huso muscular (receptores de estiramiento dinámico). Equivalen a las fibras Aα.
- Grupo Ib (o Iβ): ligeramente más delgadas y lentas. Proceden del órgano tendinoso de Golgi, que detecta la tensión muscular. También equivalen a Aα, en el rango inferior del subtipo.
Las fibras del grupo II (12-13 μm, <70 m/s) recogen información de las terminaciones en ramillete del huso muscular (receptores estáticos) y de mecanorreceptores cutáneos. Equivalen a las fibras Aβ.
Las fibras del grupo III (~5 μm, <30 m/s) son fibras finas mielínicas que transmiten dolor muscular, presión intensa y frío. Equivalen a las fibras Aδ.
Las fibras del grupo IV son amielínicas, las más lentas, y transmiten dolor sordo, calor y señales metabólicas musculares. Equivalen a las fibras C.
| Lloyd-Hunt | Erlanger-Gasser | Diámetro (μm) | Velocidad (m/s) | Receptor de origen |
|---|---|---|---|---|
| Ia | Aα | ~20 | ~120 | Terminación anuloespiral del huso (dinámica) |
| Ib | Aα (rango inferior) | 16–20 | 70–120 | Órgano tendinoso de Golgi |
| II | Aβ | 12–13 | <70 | Terminación en ramillete del huso (estática); mecanorreceptores cutáneos |
| III | Aδ | ~5 | <30 | Dolor muscular agudo; frío; presión intensa |
| IV | C | 0,5–2 | 0,5–2 | Dolor sordo; calor; señales metabólicas |
En examen, la pregunta más frecuente sobre clasificación de fibras es la correlación entre las dos nomenclaturas. Fíjate en los extremos: Ia = Aα (las más rápidas) y grupo IV = C (las más lentas y amielínicas). Los intermedios se deducen por orden.
12.2.3. Implicaciones funcionales de la clasificación
Conocer los tipos de fibras no es un ejercicio de nomenclatura. Las diferencias de diámetro, mielina y velocidad tienen consecuencias funcionales directas que explican cómo el sistema nervioso procesa la información de forma eficiente.
Principio de la línea marcada
Un potencial de acción no lleva inscrita su modalidad sensorial. Lo que determina si una señal se percibe como dolor, tacto, temperatura o sonido no es la naturaleza del potencial en sí, sino la vía anatómica y funcional por la que viaja y la corteza en la que termina.
Si se activa artificialmente una fibra táctil, la sensación percibida será tacto aunque el estímulo haya sido eléctrico. Si se estimula una vía dolorosa, el sistema nervioso interpretará la señal como dolor independientemente de cómo se haya generado. La especificidad modal depende de la conexión entre receptor periférico, fibra aferente, vías centrales y área cortical correspondiente.
Codificación de la intensidad
Un potencial de acción individual sigue la ley del todo o nada: su amplitud no varía con la intensidad del estímulo. Sin embargo, la sensación sí puede percibirse como más o menos intensa. Esto ocurre por dos mecanismos que actúan en paralelo:
Por un lado, aumenta la frecuencia de descarga en cada fibra: un estímulo más intenso genera más potenciales de acción por unidad de tiempo en la misma fibra.
Por otro, se produce reclutamiento: un estímulo débil activa pocas fibras, mientras que uno fuerte activa un número mayor. La suma de la actividad de más fibras se interpreta centralmente como una señal de mayor intensidad.
La intensidad de un estímulo no se codifica en la amplitud del potencial de acción, que es siempre la misma en una fibra dada. Se codifica en la frecuencia de disparo y en el número de fibras reclutadas. Estos dos parámetros son los que el sistema nervioso central lee para interpretar cuánto.
Velocidad y función
La velocidad de conducción no es un dato aislado: determina para qué sirve cada fibra.
Las fibras rápidas son adecuadas para funciones que exigen precisión temporal: propiocepción muscular, reflejos espinales de latencia corta, tacto discriminativo y control motor somático fino. Una motoneurona alfa que tarda demasiado en llegar al músculo haría imposible el ajuste postural rápido.
Las fibras lentas son adecuadas para señales que no requieren localización precisa ni respuesta inmediata: dolor persistente, prurito, temperatura ambiental, regulación autonómica. La velocidad baja no es un defecto, es una adaptación al tipo de información que transportan.
Dolor rápido y dolor lento
El ejemplo más claro de cómo la velocidad de conducción determina la experiencia funcional es la doble sensación dolorosa ante un estímulo nocivo intenso.
El dolor rápido aparece de forma casi inmediata: es agudo, bien localizado y fácil de describir. Lo transmiten las fibras Aδ, que con su fina mielina alcanzan velocidades de hasta 30 m/s. Su función es de alerta: informa rápidamente de que algo ha causado daño.
El dolor lento aparece después, es más persistente, difuso y se describe habitualmente como sordo, urente o mal localizado. Lo transmiten las fibras C, amielínicas y lentas (0,5-2 m/s). Su función es más relacionada con la protección prolongada de la zona dañada y con los mecanismos de sensibilización central.
La distinción entre dolor rápido (Aδ) y dolor lento (C) tiene implicaciones anestésicas. Los anestésicos locales bloquean las fibras en orden inverso a su diámetro: primero las más finas (C y Aδ), luego las intermedias y finalmente las gruesas. Esto explica por qué con una concentración anestésica baja se puede abolir el dolor sin afectar el tacto ni la propiocepción: las fibras Aβ, más gruesas, siguen conduciendo.
12.3. Transporte axonal
El axón puede medir desde unos pocos milímetros hasta más de un metro, como en el caso del nervio ciático. El soma neuronal sintetiza la práctica totalidad de las proteínas que la neurona necesita, pero el terminal sináptico está muy lejos. Para mantener la homeostasis del terminal y permitir la comunicación entre soma y periferia, existe el transporte axonal: un sistema de tráfico bidireccional activo que recorre el axón a lo largo de los microtúbulos.
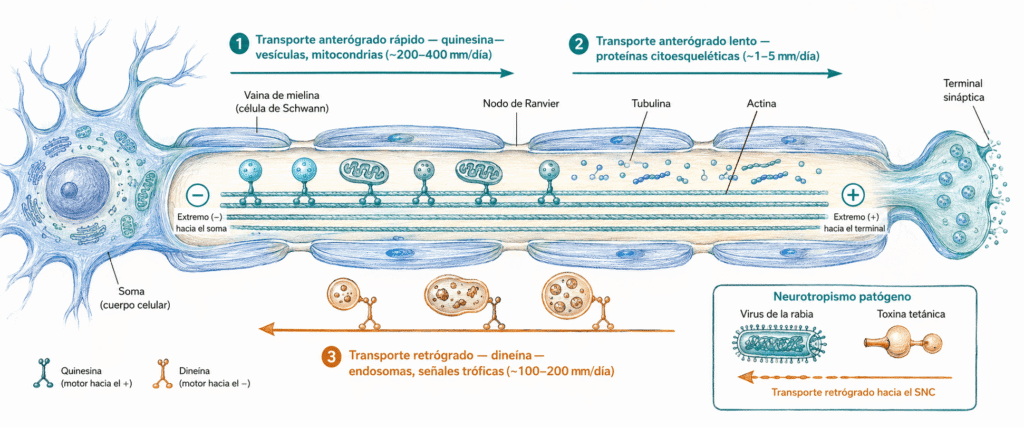
Transporte axonal: desplazamiento activo de orgánulos, vesículas, proteínas y señales a lo largo del axón mediante motores moleculares asociados a microtúbulos. Puede ser anterógrado (soma → terminal) o retrógrado (terminal → soma).
12.3.1. Transporte anterógrado
Lleva material desde el soma hacia el terminal sináptico. Existen dos modalidades según la velocidad:
- El transporte rápido (~200-400 mm/día) desplaza vesículas sinápticas, precursores de neurotransmisores, mitocondrias y componentes de membrana. El motor molecular implicado es la quinesina, que camina hacia el extremo positivo (+) de los microtúbulos, que en el axón apunta hacia el terminal.
- El transporte lento (0,1-5 mm/día) transporta proteínas citoesqueléticas solubles: tubulina, actina, neurofilamentos y enzimas del metabolismo. No viaja en vesículas, sino como masa proteica que se desplaza lentamente. Es el mecanismo por el que se renueva el armazón estructural del axón.
12.3.2. Transporte retrógrado
Lleva material desde el terminal sináptico de vuelta al soma (~100-200 mm/día). El motor implicado es la dineína, que se desplaza hacia el extremo negativo (−) de los microtúbulos, situado en el soma.
Este transporte cumple dos funciones principales. Por un lado, recicla membranas y orgánulos envejecidos que serán degradados en lisosomas del soma. Por otro, transporta señales tróficas captadas en el terminal, principalmente factores neurotróficos como el NGF (factor de crecimiento nervioso), cuya unión al receptor en la periferia genera una señal endosomal que viaja hasta el núcleo para regular la expresión génica.
Varios patógenos explotan el transporte retrógrado para acceder al SNC. La toxina tetánica y el virus de la rabia se unen a receptores en la placa neuromuscular y viajan retrógradamente por el axón motor hasta la médula espinal o el tronco encefálico, donde ejercen su efecto. Entender este mecanismo explica por qué las heridas en partes distales (pies, manos) tienen períodos de incubación más largos para el tétanos: el axón que debe recorrer la toxina es más largo.
El transporte axonal es imprescindible para la supervivencia del terminal sináptico. Si se interrumpe, el terminal degenera incluso aunque el axón permanezca físicamente intacto. Esto ocurre, por ejemplo, con ciertos fármacos que despolimerizan microtúbulos (como los alcaloides de la vinca, usados en quimioterapia), cuya neuropatía periférica se explica precisamente por este mecanismo.
12.4. Degeneración y regeneración nerviosas
Cuando un axón se lesiona, ocurren cambios tanto en el segmento distal a la lesión como en el soma neuronal. El desenlace, regeneración o muerte neuronal, depende de la gravedad de la lesión, del tipo de neurona y del entorno en que se produce.
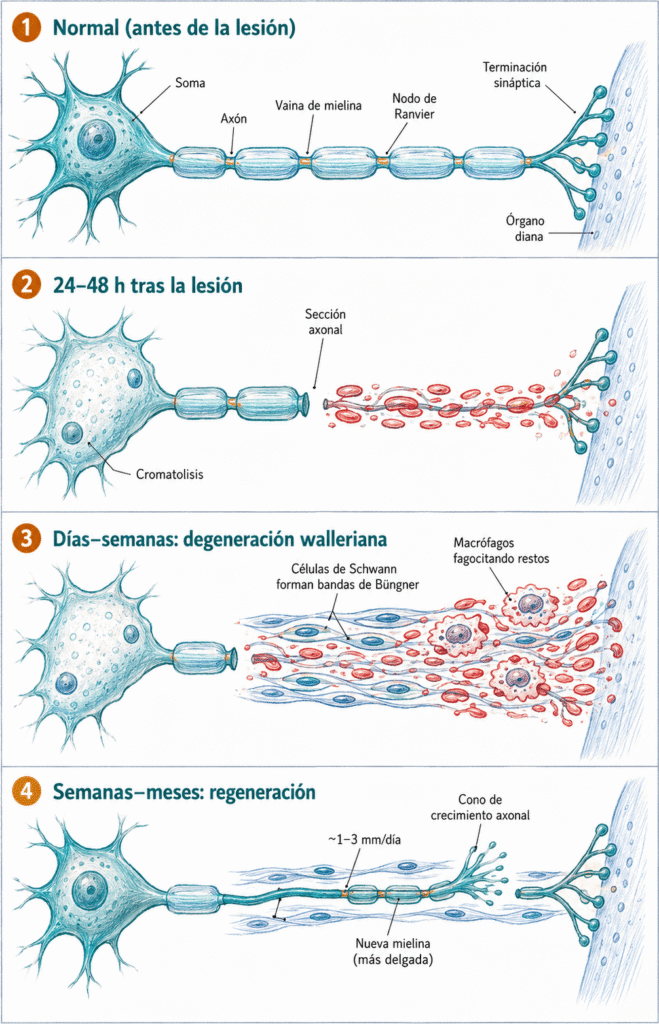
12.4.1. Degeneración walleriana
Cuando un axón se secciona, el segmento distal queda desconectado del soma y pierde el aporte de proteínas y orgánulos que dependía del transporte anterógrado. En 24-48 horas el axón distal empieza a fragmentarse; la vaina de mielina se desmenuza en ovoides lipídicos. Este proceso se denomina degeneración walleriana, descrita por Augustus Waller en 1850.
Degeneración walleriana: degeneración del segmento axonal distal a una sección nerviosa, junto con su vaina de mielina, como consecuencia de la interrupción del transporte axonal anterógrado.
Los restos del axón y la mielina son fagocitados por las células de Schwann y por macrófagos que infiltran la zona. Este proceso de limpieza es indispensable para la posterior regeneración: el territorio distal debe quedar despejado antes de que el cono de crecimiento axonal pueda avanzar.
12.4.2. Reacción axonal central
Al mismo tiempo, el soma neuronal experimenta la reacción axonal central (también llamada cromatolisis): el retículo endoplásmico rugoso se dispersa, el núcleo se desplaza a la periferia y la síntesis proteica se redirige hacia proteínas estructurales necesarias para la regeneración, en lugar de neurotransmisores.
Esta reacción es una respuesta de supervivencia y reconstrucción. Si la lesión es muy proximal o muy grave, el soma puede no sobrevivir a la desaferenciación y morir.
12.4.3. Regeneración en el SNP
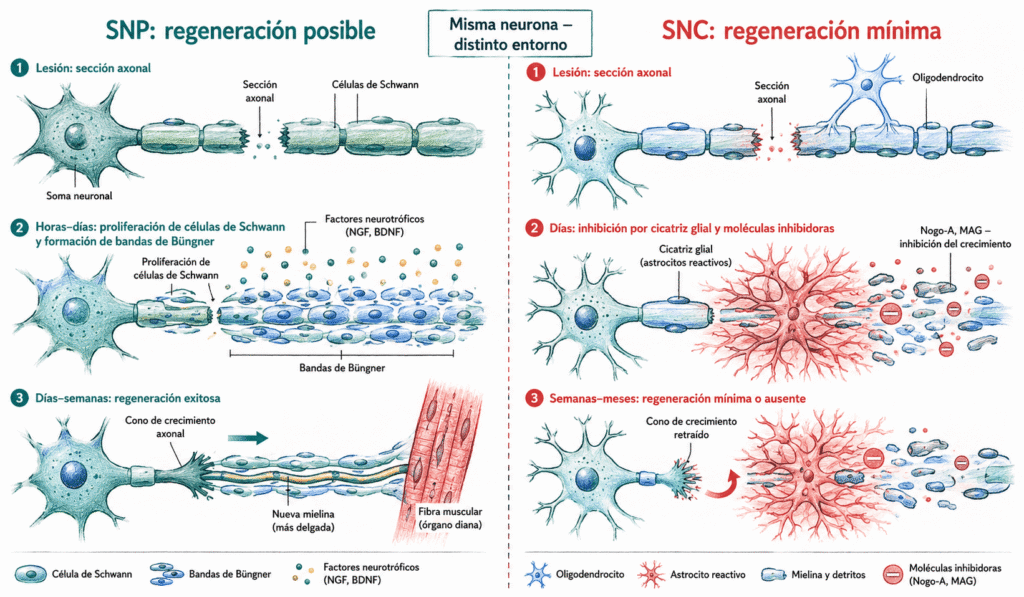
En el sistema nervioso periférico, la regeneración es posible gracias a las células de Schwann. Tras la degeneración walleriana, estas células proliferan y se alinean dentro del tubo de la lámina basal original, formando las bandas de Büngner: cordones celulares que actúan como guías físicas para el cono de crecimiento axonal.
Además, las células de Schwann secretan factores neurotróficos (NGF, BDNF, GDNF) que atraen quimiotácticamente el cono de crecimiento. El axón regenerante avanza a una velocidad de aproximadamente 1-3 mm/día. La reinervación del órgano diana no siempre es perfecta: si el axón entra en un tubo de Büngner que no corresponde a su destino original, la función recuperada puede ser incompleta o aberrante.
La velocidad de regeneración de ~1-3 mm/día tiene implicaciones pronósticas directas. Una lesión del nervio radial en el tercio medio del húmero (a unos 15-20 cm de los músculos extensores de la muñeca) puede tardar 6-8 meses en recuperar función motora. Este cálculo aproximado ayuda a establecer expectativas realistas con el paciente y a planificar el momento de la cirugía reparadora si fuera necesaria.
12.4.4. Por qué el SNC no se regenera
En el sistema nervioso central, la regeneración axonal es mínima y funcionalmente insignificante en la mayoría de los casos. Existen varias razones que se potencian mutuamente.
- En primer lugar, no hay células equivalentes a las de Schwann. Los oligodendrocitos, que producen la mielina central, no tienen capacidad de guiar el crecimiento axonal ni secretan los factores neurotróficos adecuados.
- En segundo lugar, la mielina central contiene proteínas activamente inhibidoras del crecimiento axonal, como Nogo-A, MAG (glicoproteína asociada a mielina) y OMgp, que se liberan tras la degeneración y bloquean el cono de crecimiento.
- En tercer lugar, los astrocitos reactivos forman una cicatriz glial que constituye una barrera física y química. Aunque la cicatriz protege el tejido sano circundante, impide el avance del axón en regeneración.
La neurogénesis, es decir, cómo se originan, proliferan y migran las neuronas durante el desarrollo embrionario, se estudia en Embriología. Los temas correspondientes son T12 — Desarrollo del sistema nervioso central. Vesículas encefálicas y derivados y T13 — Desarrollo del sistema nervioso periférico. Derivados de la cresta neural.
La diferencia regenerativa entre SNP y SNC no es intrínseca a la neurona, sino al entorno. Experimentos clásicos de Aguayo demostraron que neuronas del SNC pueden regenerar axones cuando se les proporciona un injerto de nervio periférico como guía: la neurona tiene la maquinaria necesaria, pero el entorno central no la deja usarla.
Un error frecuente es pensar que el SNC es incapaz de cualquier cambio tras una lesión. Lo que no hace bien es regenerar axones seccionados. Pero sí existe plasticidad sináptica: las sinapsis existentes se refuerzan, se forman nuevas colaterales de axones intactos (sprouting) y se producen reorganizaciones funcionales del mapa cortical. Esto es la base de la neurorrehabilitación, no la regeneración axonal propiamente dicha.
12.5. Plasticidad neuronal, aprendizaje y memoria
La plasticidad neuronal es la capacidad del sistema nervioso para modificar sus conexiones sinápticas en respuesta a la experiencia, el aprendizaje y la lesión.
Incluye fenómenos como el sprouting colateral de axones intactos, la potenciación a largo plazo y la reorganización de mapas corticales. No es regeneración axonal propiamente dicha, pero es el mecanismo que subyace a la neurorrehabilitación y a la memoria.
Se desarrolla en profundidad en asignaturas de Neurociencias y Fisiología del sistema nervioso de cursos superiores.
