15.1. Contexto histórico: de la teoría reticulista a la teoría neuronal
A finales del siglo XIX existía un debate fundamental sobre la organización del sistema nervioso. La posición dominante era la teoría reticulista, defendida por Camillo Golgi (la autoridad científica más influyente del momento) que concebía el sistema nervioso como una red continua de fibras interconectadas, sin separación entre células individuales.
Frente a esta visión, Santiago Ramón y Cajal defendió la teoría neuronal: el sistema nervioso era un conjunto de células individualizadas (neuronas). Cada célula tenía un principio y un final definidos y estaban separadas entre sí por espacios microscópicos. La paradoja histórica es que Cajal demostró esta teoría utilizando precisamente las técnicas de tinción desarrolladas por Golgi.
Ambos compartieron el Premio Nobel de Fisiología o Medicina en 1906, a pesar de sostener posiciones científicas opuestas. La teoría neuronal de Cajal es hoy el fundamento de toda la neurociencia.
A principios del siglo XX, Charles Sherrington introdujo el concepto de sinapsis para describir el punto de comunicación entre neuronas. En 1995 se estableció que las sinapsis son estructuras dinámicas: aparecen, se desarrollan, se estabilizan y pueden desaparecer a lo largo de la vida. Una neurona puede modificar el número y la eficacia de sus sinapsis en respuesta a la actividad, representando la base celular del aprendizaje y la memoria.
15.2. Concepto de sinapsis
Una sinapsis es la estructura especializada que permite la comunicación funcional entre una neurona y otra célula, que puede ser otra neurona, una célula muscular o una célula glandular. Es el punto donde la señal eléctrica generada en una neurona se transmite a la célula siguiente.
Unión funcional especializada entre una neurona presináptica y una célula postsináptica (neurona, músculo o glándula) que permite la transmisión de señales del sistema nervioso. Término introducido por Sherrington (1897).
La señal que llega a la presinapsis es siempre eléctrica, es decir, un potencial de acción. Lo que varía entre tipos de sinapsis es el mecanismo por el que esa señal se transmite a la célula postsináptica.
15.3. Clasificación de las sinapsis
15.3.1. Sinapsis eléctricas
En las sinapsis eléctricas, la señal se transmite directamente mediante corrientes iónicas que fluyen de una célula a la siguiente a través de canales proteicos especializados: las uniones GAP. Las membranas de ambas células están separadas por tan solo ~3 nm y unidas físicamente por estas estructuras de canal.
Sus características funcionales son:
- Bidireccionalidad: la corriente puede fluir en ambos sentidos, de la célula presináptica a la postsináptica y viceversa, aunque en algunos casos existe cierta rectificación preferente.
- Sin retardo sináptico: la transmisión es prácticamente instantánea — no hay pasos intermedios de liberación ni difusión de mensajeros.
- Sincronización: permiten que grupos de células actúen de forma coordinada y simultánea, lo que las hace ideales en tejidos que requieren contracción sincrónica.
15.3.2. Sinapsis químicas
En las sinapsis químicas no existe contacto físico entre las células. La separación entre membranas es de ~30 nm. La señal eléctrica presináptica se convierte en una señal química: el potencial de acción desencadena la liberación de una molécula mensajera denominada neurotransmisor, que difunde a través de la hendidura sináptica y se une a receptores específicos en la membrana postsináptica.
Sus características funcionales son:
- Unidireccionalidad: la señal solo puede avanzar de la presinapsis a la postsinapsis, nunca en sentido contrario, porque el neurotransmisor se libera en la presinapsis y los receptores están en la postsinapsis.
- Retardo sináptico: existe un retraso de ~0,5 ms entre la llegada del PA a la presinapsis y la respuesta postsináptica, debido al tiempo necesario para la liberación, difusión y unión del neurotransmisor.
- Versatilidad: permiten amplificación de la señal, modulación, inhibición y plasticidad sináptica, propiedades que hacen posible el procesamiento complejo de información.
| Característica | Sinapsis eléctrica | Sinapsis química |
|---|---|---|
| Separación entre membranas | ~3 nm | ~30 nm |
| Estructura de transmisión | Uniones GAP (conexones) | Neurotransmisor + receptor específico |
| Direccionalidad | Bidireccional | Unidireccional |
| Retardo sináptico | No (~0 ms) | Sí (~0,5 ms) |
| Amplificación de señal | No | Sí |
| Plasticidad | Limitada | Alta |
| Frecuencia en humanos | Poco frecuente | Predominante |
| Ejemplos | Músculo cardíaco, músculo liso, algunas conexiones SNC | Prácticamente todas las sinapsis del SNC y SNP |
Las sinapsis eléctricas sacrifican versatilidad a cambio de velocidad y sincronización. Las químicas sacrifican velocidad a cambio de modulación, amplificación y plasticidad.
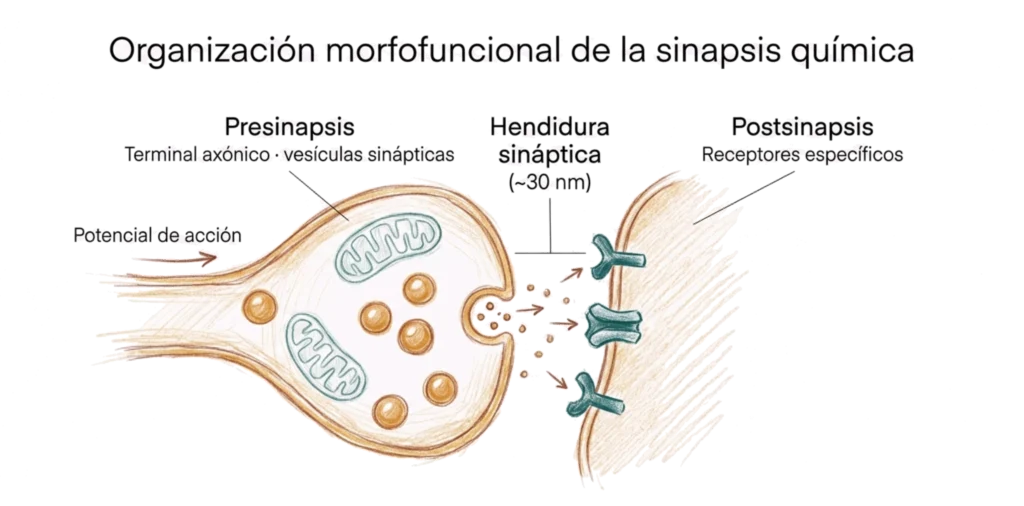
15.4. Organización morfofuncional de la sinapsis
Toda sinapsis (eléctrica o química) se organiza en tres compartimentos morfofuncionales.
15.4.1. Presinapsis
La presinapsis es el componente presináptico de la sinapsis, normalmente un terminal axónico (botón sináptico), aunque también puede ser una dendrita, un soma o incluso un axón en tránsito. Contiene la maquinaria necesaria para transmitir la señal: en las sinapsis eléctricas, los hemiconexones; en las químicas, las vesículas sinápticas cargadas de neurotransmisor.
15.4.2. Hendidura sináptica
La hendidura sináptica es el espacio extracelular entre la membrana presináptica y la postsináptica. En las sinapsis eléctricas mide ~3 nm y está atravesada por los conexones. En las químicas mide ~30 nm y es el espacio por el que difunde el neurotransmisor.
15.4.3. Postsinapsis
La postsinapsis es la membrana de la célula receptora de la señal. En las sinapsis eléctricas contiene los hemiconexones que completan el canal. En las químicas contiene los receptores específicos para el neurotransmisor liberado desde la presinapsis.
15.5. Sinapsis eléctricas: estructura y funcionamiento
15.5.1. Uniones GAP y conexones
La estructura que hace posible la sinapsis eléctrica es la unión GAP (gap junction o unión en hendidura). Es una agrupación de canales proteicos que conectan directamente el citoplasma de dos células adyacentes, permitiendo el paso de iones y moléculas pequeñas.
Cada canal individual se denomina conexón y está formado por dos hemiconexones enfrentados, uno aportado por cada célula. Cada hemiconexón está compuesto por seis subunidades proteicas denominadas conexinas, proteínas transmembrana de ~25 kDa que se disponen simétricamente alrededor de un poro central.
Las conexinas pueden ser de distintos tipos según el tejido. Se han identificado más de 20 isoformas en mamíferos, codificadas por genes distintos. La combinación de conexinas determina las propiedades biofísicas del canal.
El poro del conexón tiene un diámetro suficiente para dejar pasar no solo iones sino también moléculas de hasta ~1500 Da, incluyendo segundos mensajeros como el AMPc, el IP₃ y el Ca²⁺. Esta permeabilidad hace que las uniones GAP sean también vías de comunicación metabólica entre células, no solo eléctrica.
Canal intercelular formado por dos hemiconexones enfrentados, cada uno compuesto por seis conexinas. Permite el paso de iones y moléculas de hasta ~1500 Da entre células adyacentes. Unidad estructural de las uniones GAP.
15.5.2. Regulación del conexón
El conexón no es un canal permanentemente abierto, sino que puede cerrarse en respuesta a determinadas señales:
- El Ca²⁺ intracelular provoca el cierre del conexón cuando su concentración aumenta. Esto ocurre típicamente en situaciones de daño celular: un aumento de Ca²⁺ intracelular señala lesión, y el cierre del conexón aísla la célula dañada del resto del tejido, evitando la propagación del daño.
- Los iones H⁺ (descenso del pH intracelular) también inducen el cierre. La acidosis intracelular es otro marcador de estrés o daño celular.
Ambos mecanismos funcionan como sistemas de protección: la célula dañada se desconecta del tejido sano cerrando sus conexones.
Ante daño celular → ↑ Ca²⁺ intracelular y ↓ pH → cierre del conexón → aislamiento de la célula dañada → protección del tejido circundante.
15.5.3. Distribución fisiológica
Las sinapsis eléctricas son poco frecuentes en el sistema nervioso humano en comparación con las químicas. Se encuentran principalmente en:
- Músculo cardíaco: los discos intercalares entre cardiomiocitos contienen abundantes uniones GAP que sincronizan la contracción del miocardio. Son fundamentales para el funcionamiento coordinado del corazón como bomba.
- Músculo liso visceral: las uniones GAP entre células musculares lisas permiten la propagación de ondas de contracción coordinadas en órganos como el intestino y el útero.
- Algunas conexiones del SNC: determinados circuitos del sistema nervioso central utilizan sinapsis eléctricas para sincronización rápida, como en ciertos núcleos del tronco del encéfalo y en las células amacrinas de la retina.
- Organismos invertebrados y peces: en estos animales las sinapsis eléctricas son mucho más frecuentes que en mamíferos, donde predominan las sinapsis químicas.
15.6. Tipos de sinapsis según su localización
La posición de la sinapsis sobre la neurona postsináptica no es arbitraria — determina su influencia sobre el disparo del potencial de acción y su función en el circuito neuronal. Se distinguen cinco tipos según los compartimentos celulares que conectan:
- Las sinapsis axo-dendríticas son las más frecuentes. El terminal axónico presináptico contacta con una dendrita de la neurona postsináptica. El contacto puede establecerse sobre la espina dendrítica (pequeñas excrecencias que aumentan enormemente la superficie de contacto y son el sustrato morfológico de la plasticidad sináptica) o sobre el tronco dendrítico. Dos neuronas pueden compartir un gran número de sinapsis axo-dendríticas simultáneamente. Suelen ser sinapsis excitadoras.
- Las sinapsis axo-somáticas contactan directamente sobre el soma de la neurona postsináptica. Por su proximidad al cono axónico tienen una influencia considerable sobre el disparo. Suelen ser inhibidoras. Activan canales de Cl⁻ que introducen cargas negativas en el soma y dificultan la generación del potencial de acción.
- Las sinapsis axo-axónicas se establecen entre el terminal axónico de una neurona y el axón de otra, frecuentemente sobre el propio terminal presináptico. Son el sustrato morfológico de la modulación presináptica: al actuar directamente sobre el terminal presináptico regulan la cantidad de neurotransmisor liberado sin alterar la frecuencia de los potenciales de acción de la neurona modulada.
- Las sinapsis soma-somáticas y las sinapsis dendro-dendríticas son poco frecuentes. Las dendro-dendríticas suelen ser excitadoras por el tipo de canal iónico predominante en la membrana dendrítica.
La localización de una sinapsis sobre la neurona postsináptica determina su peso en la integración sináptica. Las sinapsis más próximas al cono axónico (axo-somáticas) tienen mayor influencia que las distales (axo-dendríticas en espinas lejanas), cuyos potenciales se atenúan más al propagarse pasivamente hasta el punto de integración.
Las sinapsis químicas — mecanismo de liberación de neurotransmisor, receptores y potenciales postsinápticos — se desarrollan en T16 — Características generales de las sinapsis químicas.
