18.1. Generalidades y localización de las sinapsis catecolaminérgicas
Las catecolaminas forman un grupo de tres neurotransmisores con estructura química compartida: noradrenalina (norepinefrina), adrenalina (epinefrina) y dopamina. Pertenecen a la familia de las aminas biógenas, dentro de los neurotransmisores de bajo peso molecular. La adrenalina y la noradrenalina actúan además como hormonas circulantes, sintetizadas por la médula suprarrenal (80% adrenalina, 20% noradrenalina).
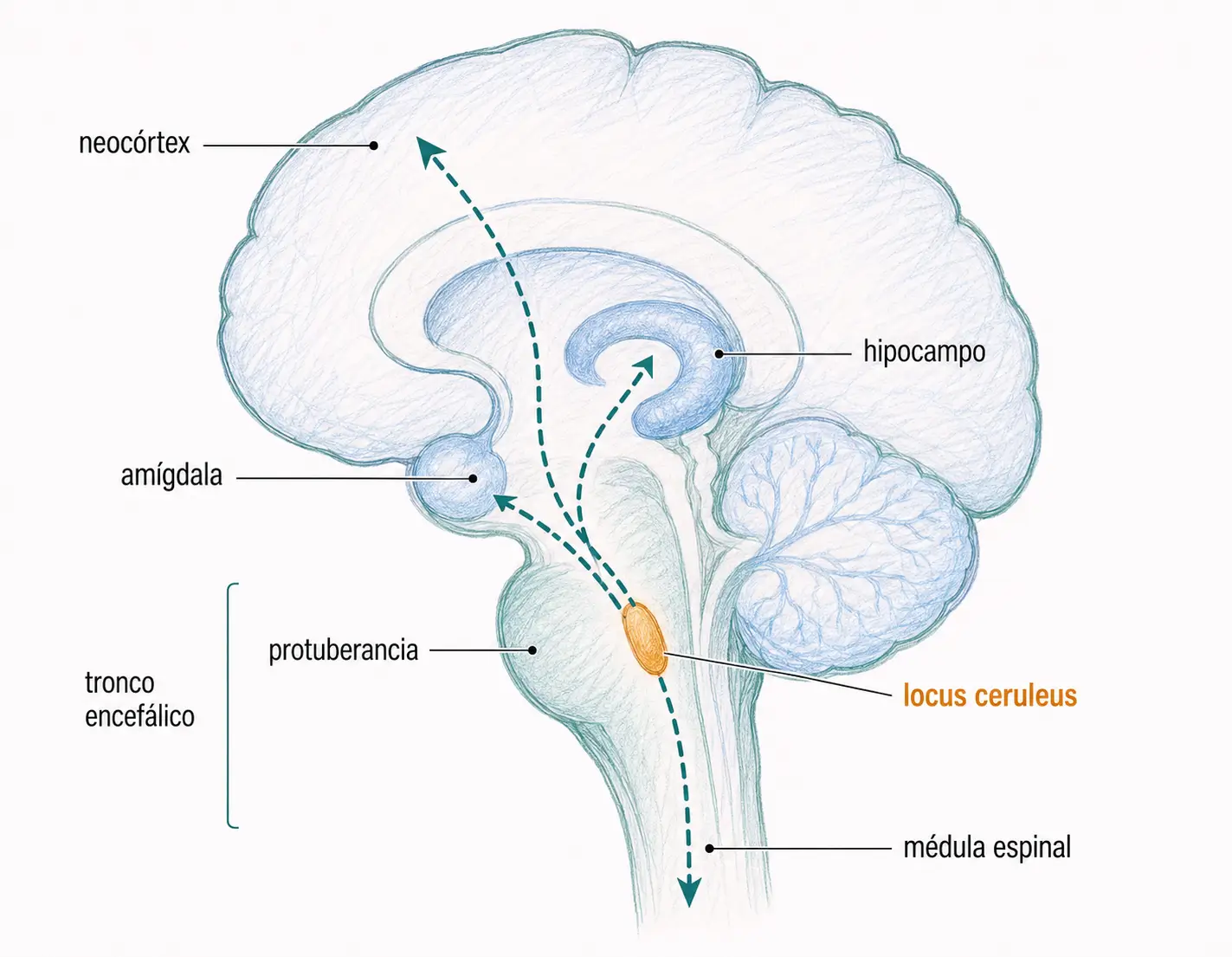
Los somas de las neuronas catecolaminérgicas se concentran principalmente en el tronco encefálico y se proyectan ampliamente al cerebro anterior.
- Las neuronas noradrenérgicas se agrupan en el locus ceruleus (protuberancia), desde donde se proyectan al mesencéfalo y el telencéfalo. La noradrenalina es también el principal neurotransmisor postganglionar del sistema nervioso simpático.
- Las neuronas adrenérgicas se localizan en el bulbo rostral. Su función en el SNC es poco conocida; el papel principal de la adrenalina es hormonal.
- Las neuronas dopaminérgicas se concentran en la sustancia negra y en el área tegmental ventral, con proyecciones hacia el estriado y el sistema límbico.
Las neuronas catecolaminérgicas tienen una frecuencia basal de descarga muy lenta, pero responden a estímulos con ráfagas de potenciales de acción que aumentan transitoriamente la liberación del neurotransmisor.
18.2. Funciones fisiológicas de las catecolaminas
- Noradrenalina: participa en procesos de atención, aprendizaje, respuesta y ansiedad, con acciones predominantes en el neocórtex, el hipocampo y la amígdala. Las neuronas del locus ceruleus se activan en situaciones de estrés, facilitando la vigilancia y la respuesta adaptativa.
- Adrenalina: prepara al organismo para la respuesta de lucha o huida, incrementando la frecuencia y la fuerza cardíaca y acelerando el metabolismo. Actúa activando el sistema nervioso simpático.
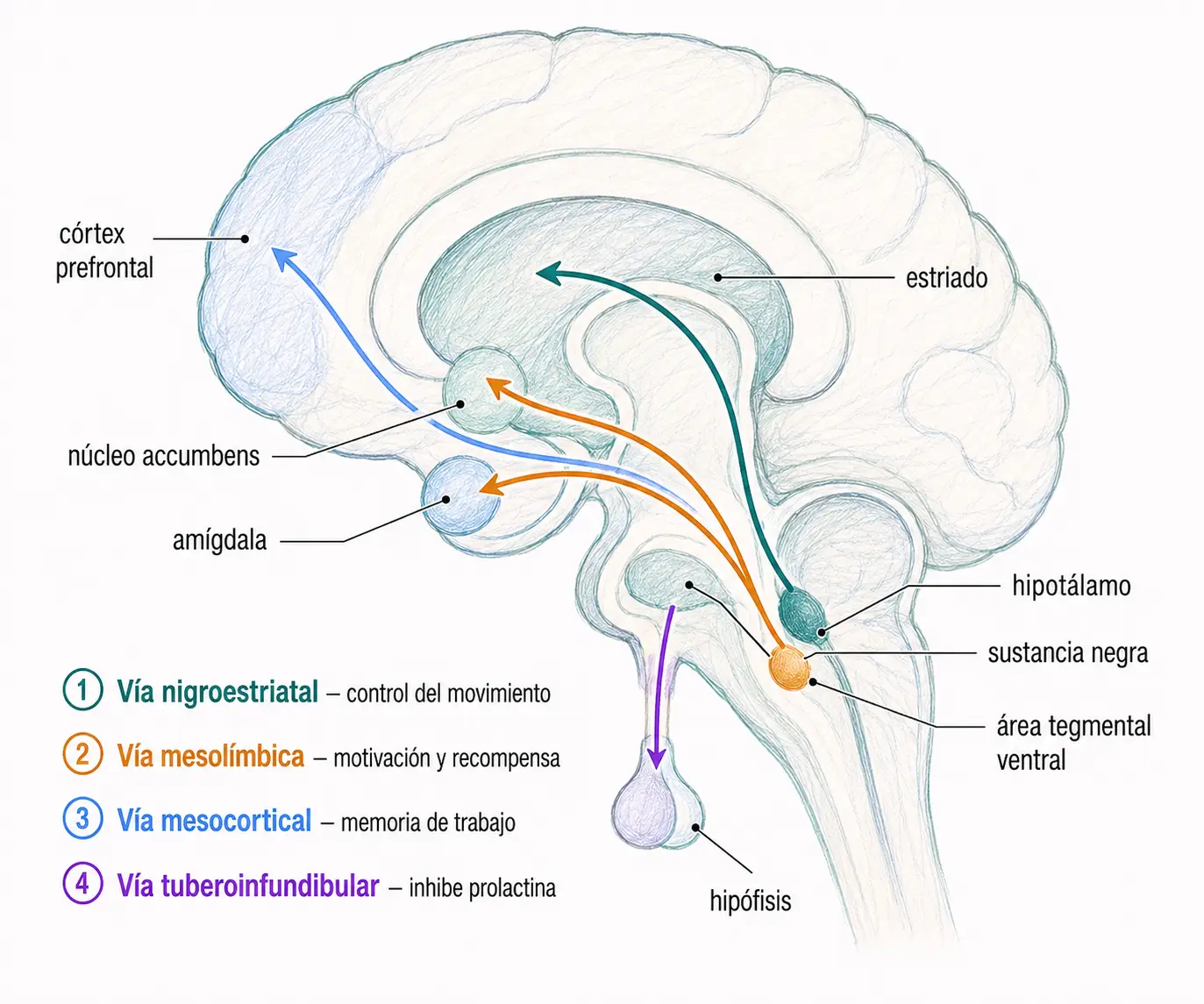
- Dopamina: interviene en cuatro vías principales, cada una con función y relevancia clínica propias:
- Nigroestriatal.
- Mesolímbica.
- Mesocortical.
- Tuberoinfundibular.
| Vía | Origen | Destino | Función principal | Consecuencia del bloqueo D2 |
|---|---|---|---|---|
| Nigroestriatal | Sustancia negra | Estriado | Control del movimiento involuntario | Parkinsonismo farmacológico |
| Mesolímbica | Área tegmental ventral | Núcleo accumbens, amígdala | Motivación, recompensa, emoción | Reducción de síntomas positivos en esquizofrenia |
| Mesocortical | Área tegmental ventral | Córtex prefrontal | Memoria de trabajo, control ejecutivo | Síntomas negativos / déficit cognitivo |
| Tuberoinfundibular | Hipotálamo (núcleo arcuato) | Hipófisis anterior | Inhibición tónica de prolactina | Hiperprolactinemia (galactorrea, amenorrea) |
La enfermedad de Parkinson se produce por la pérdida masiva y progresiva de neuronas dopaminérgicas de la sustancia negra. La depleción de dopamina en el estriado produce los síntomas cardinales: rigidez, hipertonía, temblor de reposo y bradicinesia. La afectación de la vía mesolímbica explica la depresión frecuentemente asociada.
Fármacos como la cocaína, las anfetaminas y el éxtasis aumentan la dopamina extrasináptica bloqueando su recaptación o aumentando su liberación, lo que explica sus efectos euforizantes y el potencial adictivo.
Los antipsicóticos clásicos (haloperidol, clorpromazina) son antagonistas D2 no selectivos. Su eficacia en la esquizofrenia se explica por el bloqueo de la hiperdopaminergia mesolímbica (síntomas positivos). Sus efectos secundarios extrapiramidales (parkinsonismo farmacológico) se deben al bloqueo D2 en la vía nigroestriatal. La hiperprolactinemia (galactorrea, amenorrea) se produce por bloqueo D2 en la vía tuberoinfundibular. Los antipsicóticos atípicos reducen estos efectos adversos por su menor afinidad D2 nigroestriatal.
18.3. Síntesis y almacenamiento de catecolaminas
Las tres catecolaminas se sintetizan a partir de un precursor común: la L-tirosina, un aminoácido esencial. La ruta es secuencial y cada paso añade una modificación química:
| Paso | Sustrato → Producto | Enzima | Distribución |
|---|---|---|---|
| 1 | Tirosina → L-DOPA | Tirosina hidroxilasa | Todas las neuronas catecolaminérgicas. Paso limitante de la síntesis. |
| 2 | L-DOPA → Dopamina | DOPA descarboxilasa | Muchos tipos celulares. También cataliza Trp → Serotonina. |
| 3 | Dopamina → Noradrenalina | Dopamina-β-hidroxilasa | Interior de las vesículas sinápticas. Neuronas noradrenérgicas y células cromafines. |
| 4 | Noradrenalina → Adrenalina | Feniletanolamina-N-metiltransferasa (PNMT) | Neuronas adrenérgicas y células cromafines de la médula suprarrenal. |
La dopamina-β-hidroxilasa está localizada en el interior de las vesículas sinápticas, no en el citosol. Esto significa que el paso dopamina → noradrenalina ocurre dentro de la vesícula, una vez que la dopamina ya ha sido empaquetada. Las neuronas que solo expresan hasta el paso 2 liberan dopamina; las que expresan la enzima vesicular hasta el paso 3 liberan noradrenalina.
Una vez sintetizadas, las catecolaminas se almacenan en vesículas sinápticas mediante un transportador vesicular de monoaminas (VMAT) dependiente de ATP, que trabaja contra gradiente de concentración (concentración vesicular ~0,5 M frente a ~0,1 nM en el citosol).
La reserpina bloquea el VMAT, impidiendo la entrada de catecolaminas en las vesículas. Sin protección vesicular, las catecolaminas son degradadas rápidamente por la MAO citoplasmática. Fue uno de los primeros fármacos antihipertensivos; hoy tiene uso limitado por sus efectos secundarios depresivos.
18.4. Liberación de catecolaminas
La llegada del potencial de acción al terminal presináptico abre canales de Ca²⁺ voltaje-dependientes y desencadena la exocitosis de las vesículas, con el mismo mecanismo SNARE descrito en el Tema 16: Características generales de las sinapsis químicas.
Las aminas simpatomiméticas, como la tiramina, pueden desplazar catecolaminas fuera de las vesículas al interior del citosol, provocando su liberación masiva no vesicular. El resultado es un aumento brusco de catecolaminas en la hendidura sináptica y en sangre, con elevación intensa de la presión arterial.
18.5. Receptores adrenérgicos
Todos los receptores adrenérgicos son metabotrópicos (GPCRs): no se ha identificado ningún receptor adrenérgico ionotrópico. Se clasifican en dos grandes familias, α y β, con los siguientes subtipos:
| Familia | Subtipos | 2.º mensajero | Antagonista prototípico |
|---|---|---|---|
| α1 | α1A, α1B, α1D | ↑ IP3/DAG (hidrólisis de fosfoinositol) | Fentolamina, prazosina |
| α2 | α2A, α2B, α2C | ↓ adenilato ciclasa → ↓ AMPc | Yohimbina |
| β | β1, β2, β3 | ↑ adenilato ciclasa → ↑ AMPc | Propranolol |
La adrenalina se une con afinidad similar a receptores α y β. La noradrenalina tiene mayor preferencia por los receptores α. El efecto final sobre un órgano depende del subtipo de receptor predominante en ese tejido, no del neurotransmisor en sí.
Los receptores α2 actúan también como autorreceptores presinápticos: cuando detectan la acumulación de noradrenalina en la hendidura, inhiben la fusión de vesículas y reducen la síntesis del neurotransmisor, constituyendo un mecanismo de retroalimentación negativa.
El receptor β3 tiene una distribución diferente a β1 y β2: predomina en el tejido adiposo marrón, donde media la termogénesis, y en el músculo detrusor de la vejiga. El mirabegrón, agonista β3 selectivo, se usa en el tratamiento de la vejiga hiperactiva precisamente por este perfil de distribución.
18.6. Receptores dopaminérgicos
Los receptores de dopamina se dividen en dos familias:
- Familia D1 (D1 y D5): activan la adenilato ciclasa → ↑ AMPc.
- Familia D2 (D2, D3 y D4): inhiben la adenilato ciclasa → ↓ AMPc.
Al igual que los α2 adrenérgicos, los receptores D2 actúan como autorreceptores presinápticos en las sinapsis dopaminérgicas, reduciendo la frecuencia de descarga y la síntesis y liberación de dopamina.
18.7. Recaptación y degradación
La terminación de la señal catecolaminérgica ocurre por tres mecanismos:
- Autorreceptores presinápticos (α2 y D2): inhiben directamente la síntesis y la exocitosis antes de que el neurotransmisor alcance la postsinapsis, regulando la señal en origen.
- Recaptación: es el mecanismo principal. Transportadores de membrana específicos, dependientes de Na⁺, reintroducen las catecolaminas en el terminal presináptico para su reutilización. La noradrenalina y la adrenalina son recaptadas por el NET (norepinephrine transporter, SLC6A2); la dopamina lo es por el DAT (dopamine transporter, SLC6A3). Esta distinción es farmacológicamente relevante: el metilfenidato bloquea preferentemente DAT, mientras que la duloxetina actúa principalmente sobre NET.
- Degradación enzimática intracelular: las catecolaminas recaptadas o no empaquetadas en vesículas son degradadas por dos enzimas presentes en neuronas y células gliales del SNC:
| Enzima | Localización | Sustratos principales | Inhibidor |
|---|---|---|---|
| MAO-A | Membrana externa mitocondrial | Noradrenalina, serotonina, tiramina | Clorgilina |
| MAO-B | Membrana externa mitocondrial | Dopamina, β-feniletamina | Deprenilo (selegilina) |
| COMT | Citosol y espacio extracelular | Todas las catecolaminas | Entacapona, tolcapona |
Los antidepresivos tricíclicos (imipramina, amitriptilina) bloquean de forma no selectiva NET y el transportador de serotonina (SERT), amplificando la señal noradrenérgica y serotoninérgica. Los inhibidores de recaptación de serotonina-noradrenalina (IRSN), como la venlafaxina o la duloxetina, tienen un perfil más selectivo sobre NET y SERT con menor bloqueo de otros receptores, lo que mejora su tolerabilidad.
La MAO solo actúa sobre catecolaminas libres en el citosol, no sobre las almacenadas en vesículas. La reserpina explota esta vulnerabilidad: al vaciar las vesículas, expone el neurotransmisor a la degradación por MAO.
La COMT utiliza S-adenosilmetionina (SAM) como cosustrato donador de grupos metilo. Actúa tanto en el citosol neuronal como en el espacio extracelular, siendo la principal vía de degradación periférica.
Los productos finales de degradación son medibles en orina y tienen valor diagnóstico. La degradación de noradrenalina genera ácido vanililmandélico (VMA) y DHPG (dihidroxifenilglicol). La degradación de dopamina genera ácido homovanílico (HVA). La elevación de estos metabolitos en orina de 24 horas es característica del feocromocitoma (tumor de la médula suprarrenal) y del neuroblastoma.
Síndrome del queso: los inhibidores de la MAO-A (IMAOs) usados como antidepresivos impiden la degradación de la tiramina dietética (presente en queso curado, vino tinto, frutos secos). La tiramina acumulada desplaza catecolaminas de las vesículas, provocando una crisis hipertensiva potencialmente mortal. La interacción no se identificó hasta los años 60, cuando se produjeron muertes en pacientes tratados con IMAOs sin restricción dietética. Hoy es de obligado consejo en todos los pacientes con estos fármacos.
