28.1. Qué es un receptor sensorial
El sistema nervioso no tiene acceso directo al mundo. Todo lo que percibimos, desde el tacto de una superficie hasta la presión arterial en las arterias, llega a través de una misma puerta: los receptores sensoriales.
Un receptor sensorial es una estructura especializada en detectar una forma concreta de energía o de cambio en el entorno, y en convertirla en una señal eléctrica que el sistema nervioso puede procesar. Ese proceso de conversión se llama transducción sensorial.
Transducción sensorial: transformación de la energía de un estímulo en una variación del potencial de membrana (potencial de receptor). Si esa variación supera el umbral, se desencadenan potenciales de acción que viajan hacia el sistema nervioso central.
Los receptores sensoriales pueden ser tan simples como una terminación nerviosa libre (una única neurona con su extremo libre en el tejido) o tan complejos como las células ciliadas del órgano de Corti. En todos los casos, la función es la misma: leer el estímulo y traducirlo.
28.2. Clasificación de los receptores sensoriales
Existen varias formas de clasificar los receptores sensoriales, y todas se usan en la literatura. La más útil desde el punto de vista fisiológico es la que atiende a la naturaleza del estímulo que detectan.
28.2.1. Clasificación por la naturaleza del estímulo
- Mecanorreceptores: detectan energía mecánica (presión, vibración, estiramiento, sonido).
- Quimiorreceptores: detectan sustancias químicas (gusto, olfato, presión de O₂, pH sanguíneo).
- Termorreceptores: detectan cambios de temperatura (frío y calor tienen receptores distintos).
- Fotorreceptores: detectan energía lumínica. Son los receptores más especializados del organismo.
- Nociceptores: detectan estímulos potencialmente dañinos, de naturaleza mecánica, química o térmica intensa.
28.2.2. Clasificación por el origen de la información
- Exteroceptores: reciben información del exterior del organismo (piel, ojos, oídos).
- Interoceptores: reciben información del interior del organismo (vísceras, vasos sanguíneos, sistema endocrino).
- Propioceptores: informan sobre la posición y el movimiento del propio cuerpo (músculos, tendones, articulaciones).
Los teleceptores son exteroceptores que captan información a distancia: vista, olfato y oído. La distinción es clásica en los textos de fisiología, aunque no siempre se separan explícitamente del grupo de exteroceptores.
| Tipo | Estímulo adecuado | Ejemplos | Localización característica |
|---|---|---|---|
| Mecanorreceptor | Energía mecánica: presión, vibración, estiramiento, sonido | Corpúsculo de Meissner, corpúsculo de Pacini, células ciliadas del oído | Piel, músculo, tendón, oído interno |
| Quimiorreceptor | Estímulos químicos: moléculas odorantes, sustancias sapidas, O₂, CO₂, pH | Células olfatorias, papilas gustativas, cuerpo carotídeo | Mucosa nasal, lengua, carótida, aorta |
| Termorreceptor | Variaciones de temperatura (frío: 10–35 °C; calor: 30–45 °C) | Terminaciones de fibras Aδ (frío) y C (calor) | Piel, mucosas, hipotálamo |
| Fotorreceptor | Energía lumínica (fotones) | Conos (visión en color), bastones (visión escotópica) | Retina |
| Nociceptor | Estímulos potencialmente dañinos: mecánicos intensos, químicos, térmicos extremos | Terminaciones nerviosas libres (fibras Aδ y C) | Piel, vísceras, periostio, córnea |
28.3. El potencial de receptor y la transducción
Cuando un estímulo actúa sobre un receptor, desencadena una variación del potencial de membrana en la zona receptora. Esa variación se llama potencial de receptor (o potencial de generador).
El mecanismo varía según el tipo de receptor. En un mecanorreceptor de presión, la deformación mecánica de la membrana abre canales iónicos mecanosenibles, lo que provoca una entrada de sodio y la consiguiente despolarización local. En un fotorreceptor, un fotón activa una cascada de señalización intracelular (a través de la rodopsina y proteínas G) que finalmente modifica la conductancia de la membrana.
Lo esencial es que el potencial de receptor es una señal graduada: cuanto mayor es la intensidad del estímulo, mayor es la amplitud de la despolarización.
El potencial de receptor es graduado. El potencial de acción es todo-o-nada. El umbral es la frontera entre los dos: solo cuando el potencial de receptor supera el umbral crítico se desencadena el primer potencial de acción.
Si el estímulo es subumbral, la despolarización local se disipa sin generar potencial de acción y el estímulo no se percibe. Si supera el umbral, se inicia la propagación hacia el sistema nervioso central.
Conexión con T10 — El potencial de acción: el mecanismo de generación y propagación del potencial de acción, la ley del todo-o-nada y los períodos refractarios se desarrollan en el Tema 10 - El Potencial de acción
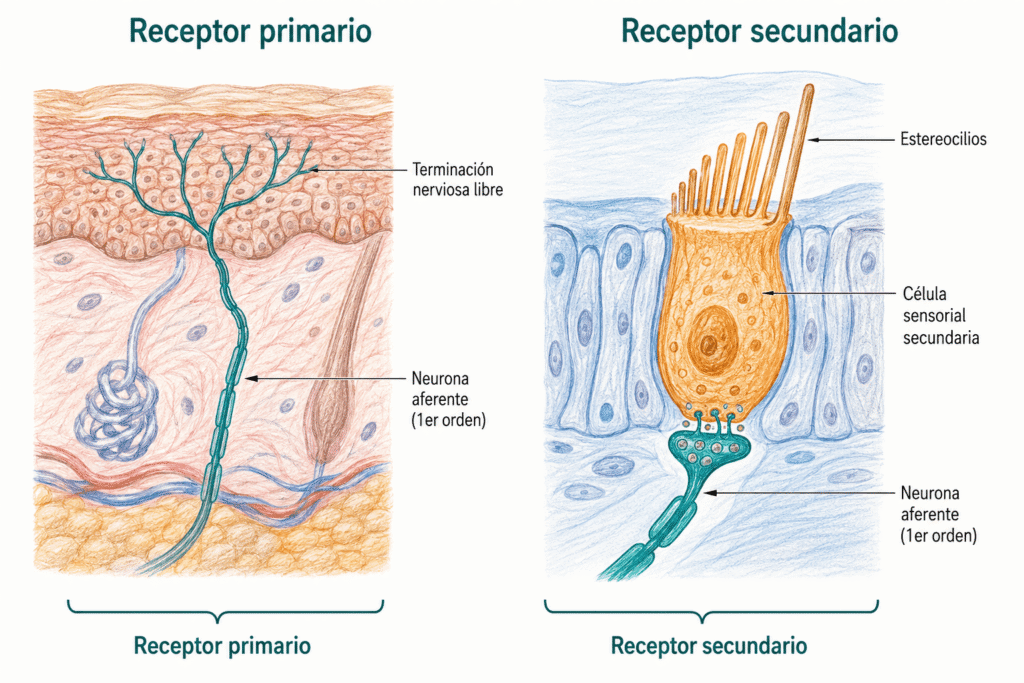
28.4. Receptores primarios y secundarios
Hay dos grandes diseños arquitectónicos para un receptor sensorial.
- En los receptores primarios, la misma célula que detecta el estímulo es una neurona: su terminación periférica actúa como zona receptora y su axón conduce los potenciales de acción hasta el sistema nervioso central. Los mecanorreceptores cutáneos y los nociceptores son receptores primarios.
- En los receptores secundarios, la detección la realiza una célula especializada que no es una neurona. Esa célula hace sinapsis con una neurona aferente, que es la que genera el potencial de acción y lo transmite. Las células ciliadas del órgano de Corti y los fotorreceptores de la retina son receptores secundarios.
Relevancia clínica: la distinción entre receptores primarios y secundarios tiene implicaciones en la fisiopatología de los sentidos especiales. Las lesiones que afectan a la célula sensorial secundaria (p. ej., pérdida de células ciliadas por ruido o ototóxicos) producen hipoacusia neurosensorial sin que la neurona aferente esté dañada.
La transducción en receptores secundarios puede ocurrir por dos vías. En algunos, el estímulo activa directamente la célula sensorial (transducción directa), como en las células ciliadas del oído, donde la deflexión de los estereocilios abre canales mecano-sensibles. En otros, la transducción es indirecta y requiere segundos mensajeros: los fotorreceptores de la retina son el ejemplo más estudiado, donde la escasa energía de un fotón se amplifica mediante la cascada de la rodopsina.
28.5. Codificación de la información sensorial
Llegar al sistema nervioso central no es suficiente: el cerebro necesita saber qué estímulo se ha detectado, dónde, con qué intensidad y durante cuánto tiempo. Esa información se codifica en los potenciales de acción antes de llegar a la corteza.
- Modalidad: viene determinada por el tipo de receptor activado y la vía nerviosa que se activa, no por la naturaleza del estímulo en sí. Un estímulo mecánico sobre un fotorreceptor se interpreta como luz (el fenómeno de los «destellos» que se ven al golpe en el ojo ilustra esto). Es la ley de las energías específicas de los nervios.
- Intensidad: se codifica de dos maneras. La primera es la frecuencia de disparo: a mayor intensidad del estímulo, mayor amplitud del potencial de receptor, más frecuencia de potenciales de acción en la neurona sensorial. La segunda es el reclutamiento espacial: los estímulos intensos activan más receptores que los débiles.
- Localización: depende del campo receptor del receptor activado. El campo receptor es la región de la superficie corporal (o del tejido) cuya estimulación activa un receptor concreto. Cuanto menor es el campo receptor y mayor la densidad de receptores en una zona, mayor es la agudeza discriminativa de esa zona.
- Duración: está ligada al fenómeno de la adaptación, que se desarrolla en la sección siguiente.
La densidad de receptores determina el umbral y la discriminación: a mayor densidad, menor umbral y mayor capacidad de localizar el estímulo con precisión. Por eso la yema de los dedos discrimina dos puntos separados 2 mm, y la espalda necesita varios centímetros.
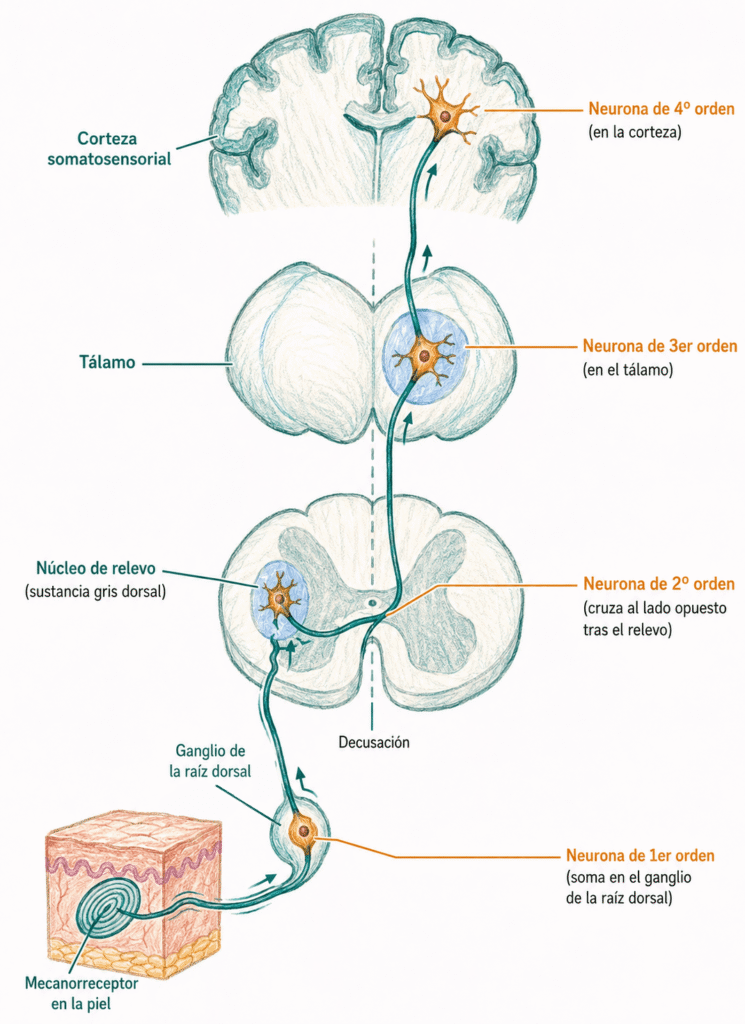
28.6. La vía sensitiva general
Los potenciales de acción generados en el receptor ascienden hasta la corteza cerebral a través de una vía formada, en general, por cuatro neuronas en serie.
- La neurona de primer orden tiene su soma en el ganglio raquídeo dorsal (para la sensibilidad somática) o en los ganglios de los pares craneales (para la sensibilidad de la cabeza). Su axón periférico llega hasta el receptor; su axón central entra en la médula espinal o el tronco encefálico.
- La neurona de segundo orden tiene su soma en los núcleos de relevo de la médula o del tronco encefálico. Sus axones cruzan la línea media y ascienden hacia el tálamo. Esta decusación es la razón por la que las lesiones en un lado del sistema nervioso central producen déficits sensitivos en el lado contralateral del cuerpo.
- La neurona de tercer orden reside en el tálamo, que actúa como estación de relevo y filtro de la información sensitiva antes de que alcance la corteza.
- La neurona de cuarto orden tiene su soma en la corteza somatosensorial. La llegada de señales a la corteza es lo que genera la consciencia del estímulo: sin activación cortical, no hay percepción consciente.
En algunas vías no existe la neurona de cuarto orden en corteza, o la vía termina antes de alcanzarla. El sistema barorreceptor, que regula la presión arterial de forma refleja, es un ejemplo: opera fundamentalmente a nivel subcortical sin generar percepción consciente.
28.7. Propiedades funcionales de los receptores
Más allá de su clasificación, los receptores sensoriales tienen propiedades que determinan cómo responden ante un estímulo real.
- Umbral: es la intensidad mínima que debe alcanzar un estímulo para generar una respuesta detectable. No todos los receptores tienen el mismo umbral: los fotorreceptores de la retina pueden detectar un único fotón en condiciones de oscuridad; los nociceptores solo se activan ante estímulos intensos.
- Especificidad: un receptor está diseñado para responder de forma preferente a un tipo de estímulo (su estímulo adecuado), aunque estímulos de otra naturaleza y suficiente intensidad también puedan activarlo. Los fotorreceptores son los más específicos del organismo; los nociceptores son los menos específicos, ya que responden a estímulos mecánicos, químicos y térmicos intensos.
- Adaptación: cuando un estímulo se mantiene constante en el tiempo, muchos receptores reducen progresivamente su tasa de disparo. Este fenómeno se llama adaptación sensorial y explica por qué dejamos de percibir el peso de la ropa o el olor de una habitación tras unos minutos de exposición.
Adaptación sensorial no es lo mismo que habituación. La adaptación ocurre en el propio receptor (disminución de la respuesta periférica). La habituación es un proceso central de aprendizaje (el sistema nervioso central "aprende" a ignorar un estímulo repetido). Son mecanismos distintos en niveles distintos.
28.8. Receptores fásicos y receptores tónicos
La adaptación no es igual en todos los receptores. Según su velocidad de adaptación, se distinguen dos grandes categorías con funciones bien diferenciadas.
- Los receptores fásicos (de adaptación rápida) se adaptan rápidamente: responden con una descarga intensa al inicio del estímulo, pero reducen o cesan su actividad si el estímulo se mantiene. Su función es señalar cambios: el inicio de un movimiento, la variación de una presión, el contacto nuevo con una superficie.
- Los receptores tónicos (de adaptación lenta) mantienen su actividad mientras dura el estímulo. Su función es informar sobre estados mantenidos: la posición de una articulación, la presión arterial sostenida, el dolor crónico.
Fásicos = detectores de cambio. Tónicos = detectores de estado.
Esta diferencia tiene implicaciones directas en cómo el sistema nervioso gestiona la información: los fásicos alertan de novedades; los tónicos mantienen el fondo de información continua sobre el estado del cuerpo.
Un mismo órgano sensorial puede contener ambos tipos. En el oído, por ejemplo, las células ciliadas externas tienen un comportamiento más fásico (ajuste de ganancia); las internas son más tónicas (transmisión de la señal auditiva).
Figura 28.1. La amplitud del potencial de receptor varía con la intensidad del estímulo (señal graduada). Si supera el umbral, se generan potenciales de acción de amplitud constante: la intensidad se codifica en frecuencia de disparo, no en amplitud.
| Característica | Receptor fásico | Receptor tónico |
|---|---|---|
| Adaptación | Rápida | Lenta o nula |
| Respuesta al estímulo mantenido | Cesa o se reduce notablemente | Se mantiene durante todo el estímulo |
| Función principal | Detectar cambios, movimiento, novedades | Informar sobre estados continuos |
| Ejemplos | Corpúsculo de Pacini (vibración), corpúsculo de Meissner (tacto superficial), receptores foliculares del pelo | Receptores musculares de estiramiento (huso neuromuscular), barorreceptores, nociceptores C |
| Fibras asociadas típicas | Aβ (mielinizadas, conducción rápida) | C (amielínicas, conducción lenta) y Aδ |
