20.1. Generalidades de las sinapsis glutamatérgicas
El glutamato es el neurotransmisor excitador más abundante del SNC en mamíferos. Las neuronas que lo sintetizan y liberan se denominan neuronas glutamatérgicas, y están presentes en prácticamente todos los circuitos del encéfalo y la médula espinal.
Neurotransmisor glutamatérgico: el glutamato (ácido glutámico) actúa como neurotransmisor excitador al unirse a receptores que abren canales catiónicos, lo que despolariza la membrana postsináptica y acerca el potencial de membrana al umbral de disparo. Su efecto excitador no es una propiedad intrínseca de la molécula, sino una consecuencia del tipo de receptor al que se une.
El aspartato comparte propiedades similares y actúa también como neurotransmisor excitador en algunas sinapsis, aunque su relevancia funcional en el SNC humano es menor que la del glutamato.
El glutamato es también un aminoácido no esencial con funciones metabólicas importantes, entre ellas su papel en el ciclo de Krebs como α-cetoglutarato. La bioquímica de estos procesos se estudia en detalle en Bioquímica y Biología Molecular. En este tema interesa exclusivamente su función como neurotransmisor.
20.2. Síntesis, ciclo glía-neurona y recaptación del glutamato
El glutamato no puede cruzar la barrera hematoencefálica (BHE) en forma libre. La neurona obtiene su pool de glutamato a través de un ciclo metabólico que implica una colaboración estrecha con las células gliales vecinas, especialmente los astrocitos.
- La glutamina circulante cruza la BHE y entra en el astrocito.
- El astrocito convierte glutamina en glutamato mediante la enzima glutaminasa y lo transfiere a la neurona presináptica.
- La neurona empaqueta el glutamato en vesículas sinápticas y lo libera por exocitosis ante la llegada de un potencial de acción.
- Tras actuar sobre los receptores postsinápticos, el glutamato liberado es recaptado desde la hendidura sináptica: principalmente por los astrocitos y, en menor medida, por la propia terminal presináptica.
- En el astrocito, el glutamato recaptado se convierte rápidamente en glutamina mediante la glutamina sintetasa, completando el ciclo y devolviendo el precursor a la neurona.
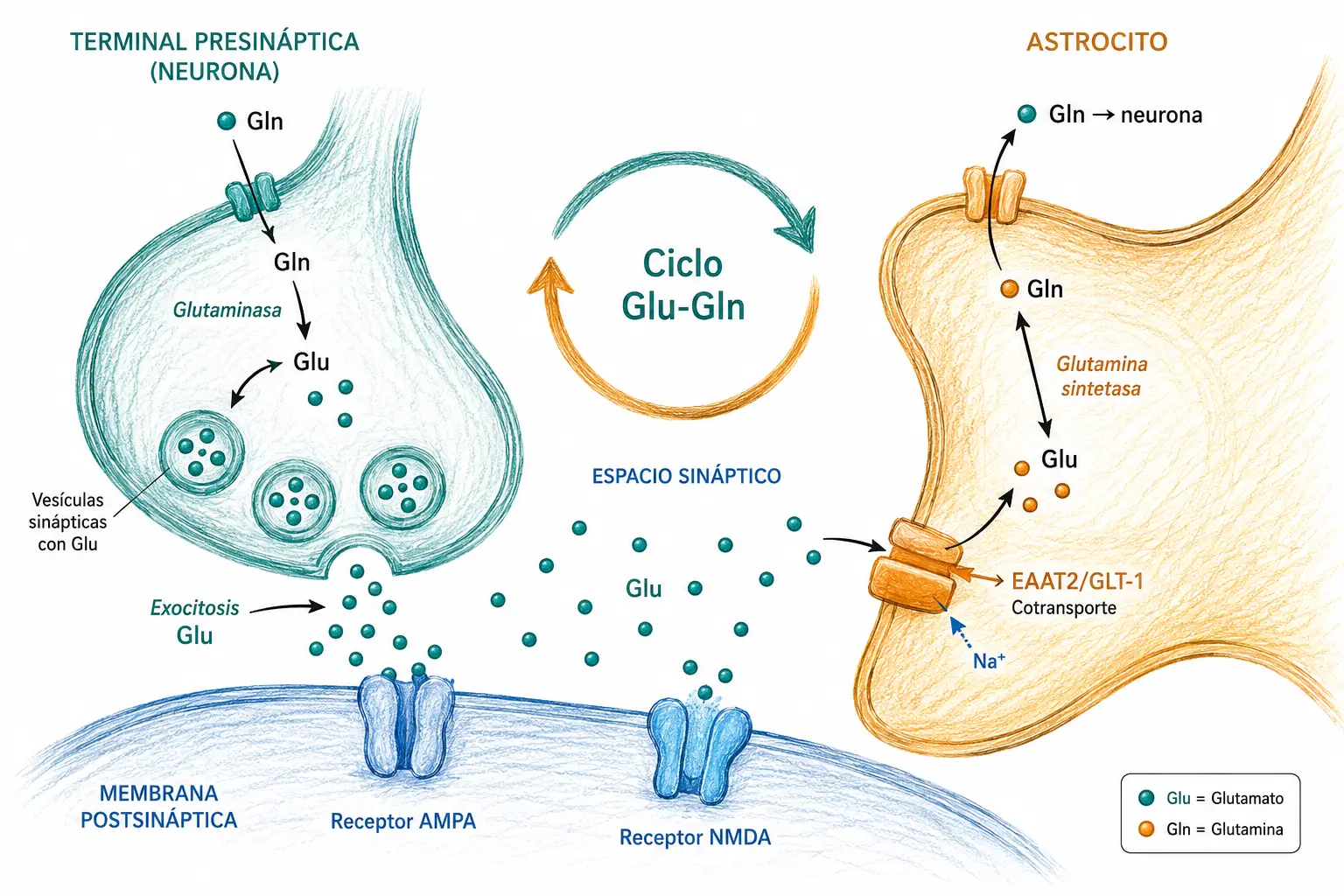
La conversión inmediata de glutamato en glutamina dentro del astrocito no es accesoria: el glutamato libre a concentraciones elevadas es tóxico para las neuronas. La glía actúa como sistema de seguridad que evita la acumulación extracelular de glutamato, protegiendo las neuronas vecinas.
20.3. Recaptación y degradación: los transportadores EAAT
La inactivación del glutamato en la hendidura sináptica no ocurre por degradación enzimática directa (a diferencia de la acetilcolina), sino fundamentalmente por recaptación activa mediante transportadores específicos.
Estos transportadores pertenecen a la familia de los transportadores de aminoácidos excitadores (EAAT, Excitatory Amino Acid Transporters). Existen cinco subtipos (EAAT1-5); el más relevante fisiológicamente es el EAAT2 (también llamado GLT-1), que se expresa mayoritariamente en los astrocitos y es responsable de aproximadamente el 90% del aclaramiento sináptico del glutamato en el cerebro.
EAAT2 es el principal mecanismo de inactivación del glutamato sináptico. Su funcionamiento correcto es esencial para prevenir la excitotoxicidad: cuando EAAT2 falla o se satura, el glutamato se acumula en la hendidura y produce daño neuronal.
Una vez dentro del astrocito, el glutamato sigue dos vías: la mayoritaria es la conversión a glutamina (ciclo descrito en el apartado anterior); una fracción menor puede entrar en el ciclo de Krebs como α-cetoglutarato tras su conversión por la aspartato aminotransferasa astrocitaria. Esta segunda vía tiene mayor relevancia metabólica que neurotransmisora.
EAAT2 en la esclerosis lateral amiotrófica (ELA): en pacientes con ELA se ha descrito una reducción significativa de la expresión de EAAT2 en la corteza motora y la médula espinal. Esta pérdida de aclaramiento glutamatérgico contribuye a la excitotoxicidad crónica de las motoneuronas. El riluzol, único fármaco aprobado hasta hace poco para la ELA, actúa en parte modulando la transmisión glutamatérgica.
20.4. Receptores ionotrópicos: AMPA, kainato y NMDA
Los receptores ionotrópicos de glutamato son canales iónicos que se abren directamente al unirse el glutamato. Se dividen en tres tipos según sus propiedades cinéticas, iónicas y farmacológicas:
| Receptor | Iones permeables | Cinética | Requisitos de activación | Función principal |
|---|---|---|---|---|
| AMPA | Na⁺, K⁺ (Ca²⁺ bajo) | Rápida | Glutamato | Transmisión excitatoria rápida |
| Kainato | Na⁺, K⁺ | Rápida | Glutamato | Transmisión excitatoria; modulación presináptica |
| NMDA | Na⁺, K⁺, Ca²⁺ (alta) | Lenta | Glutamato + coagonista + despolarización | Plasticidad sináptica (LTP), detector de coincidencia |
20.4.1. Receptores AMPA y kainato
Los receptores AMPA son los mediadores de la transmisión excitatoria rápida en la inmensa mayoría de las sinapsis glutamatérgicas del SNC. Se ensamblan como heterotetrámeros de subunidades GluA1-GluA4; la composición de subunidades determina sus propiedades biofísicas y su comportamiento de tráfico en la membrana.
Su apertura produce una despolarización de inicio rápido y corta duración. Son permeables principalmente a Na⁺ y K⁺; su permeabilidad al Ca²⁺ es generalmente baja, aunque puede aumentar en receptores que carecen de la subunidad GluA2.
Los receptores de kainato tienen una distribución más restringida y participan tanto en la transmisión postsináptica como en la modulación de la liberación presináptica de neurotransmisores.
20.4.2. Receptor NMDA: el detector de coincidencia
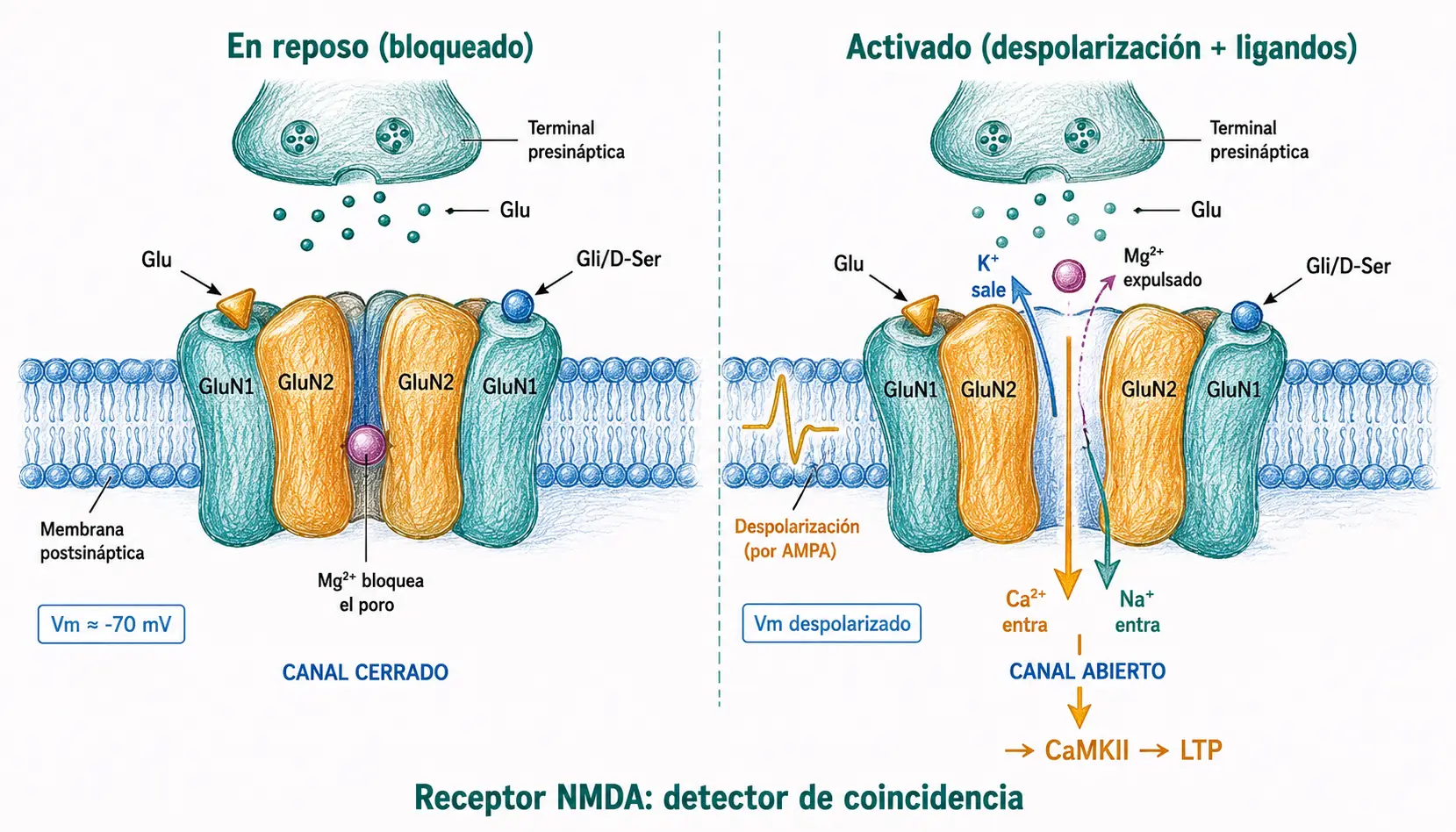
El receptor NMDA es el más complejo y biofísicamente singular de los receptores ionotrópicos de glutamato. Se ensambla como un heterotetrámero que contiene siempre dos subunidades GluN1 (que aportan el sitio de unión al coagonista) y dos subunidades GluN2 (principalmente GluN2A o GluN2B en el SNC adulto, que aportan el sitio de unión al glutamato).
Sus propiedades diferenciales son tres:
- Alta permeabilidad al Ca²⁺. A diferencia de los AMPA, los receptores NMDA son muy permeables al Ca²⁺, además de a Na⁺ y K⁺. Esta entrada de Ca²⁺ conecta la actividad sináptica con cascadas de señalización intracelular que modulan la eficacia de la sinapsis a largo plazo.
- Bloqueo voltaje-dependiente por Mg²⁺. A potenciales de membrana cercanos al reposo, un ion Mg²⁺ ocupa el poro del canal e impide el paso de iones aunque el glutamato esté unido. Solo cuando la membrana postsináptica se despolariza suficientemente, el Mg²⁺ es repelido electrostáticamente y el canal puede abrirse. Esto significa que el receptor NMDA necesita dos condiciones simultáneas para funcionar:
- La presencia de glutamato y
- Que la célula ya esté despolarizada.
- Coagonismo obligatorio. Además del glutamato, el receptor NMDA requiere la unión simultánea de un segundo ligando en un sitio distinto de la subunidad GluN1: el llamado sitio glicínico. Los dos principales ligandos endógenos de este sitio son la glicina y la D-serina.
La D-serina es sintetizada por la enzima serina racemasa en las propias neuronas glutamatérgicas, a partir de L-serina que procede en parte del metabolismo astrocitario. La evidencia actual indica que la D-serina es el coagonista predominante en los receptores NMDA sinápticos de la corteza e hipocampo, mientras que la glicina predominaría en los receptores extrasinápticos. Ambos compiten por el mismo sitio de unión.
Un error frecuente es describir la glicina como «el único coagonista» del receptor NMDA, o afirmar que «actúa como inhibidor al unirse al receptor excitador». La glicina es inhibidora cuando actúa sobre su propio receptor ionotrópico (canal de Cl⁻, bloqueado por estricnina) en médula espinal y tronco del encéfalo. En el sitio coagonista del receptor NMDA (subunidad GluN1) actúa como activador permisivo: sin ella, el canal no se abre aunque haya glutamato. Son receptores, mecanismos y contextos completamente distintos.
El receptor NMDA actúa como un detector de coincidencia: solo se activa cuando coinciden simultáneamente la presencia de glutamato en la hendidura, la unión del coagonista (glicina o D-serina) Y la despolarización suficiente de la membrana postsináptica (que retira el Mg²⁺). Este mecanismo es la base molecular de la potenciación a largo plazo y, por tanto, del aprendizaje y la memoria.
20.5. Receptores metabotrópicos de glutamato
Además de los ionotrópicos, existen receptores metabotrópicos de glutamato (mGluR), acoplados a proteína G, que modulan la eficacia sináptica de forma más lenta y duradera. Se clasifican en tres grupos:
| Grupo | Receptores | Proteína G / Efector | Efecto sobre AMPc / PLC | Localización preferente |
|---|---|---|---|---|
| I | mGluR1, mGluR5 | Gq → PLC | ↑ IP3 / DAG | Postsináptica |
| II | mGluR2, mGluR3 | Gi → adenilato ciclasa | ↓ AMPc | Pre y postsináptica |
| III | mGluR4, mGluR6, mGluR7, mGluR8 | Gi → adenilato ciclasa | ↓ AMPc | Presináptica (autorreceptores) |
Los receptores del grupo I (mGluR1 y mGluR5), situados principalmente en la membrana postsináptica, amplifican la respuesta excitatoria y contribuyen a la inducción de la LTP. Los grupos II y III actúan frecuentemente como autorreceptores presinápticos: al detectar glutamato acumulado en la hendidura, reducen la liberación de más glutamato, funcionando como mecanismo de retroalimentación negativa.
Los mecanismos de señalización intracelular vía proteína G, IP3, DAG y AMPc se estudian en detalle en el Tema 7 · Señalización intracelular.
20.6. Funciones de la transmisión glutamatérgica
El sistema glutamatérgico tiene un papel central en la fisiología del SNC que va más allá de la simple transmisión excitatoria.
- Transmisión excitatoria rápida. La función más inmediata del glutamato es mediar la comunicación sináptica excitatoria en prácticamente todos los circuitos del encéfalo: corteza cerebral, hipocampo, cerebelo, ganglios basales y médula espinal. Los receptores AMPA son los principales responsables de esta función.
- Plasticidad sináptica y memoria. A través del mecanismo NMDA-dependiente de la LTP (desarrollado en el apartado siguiente), el glutamato es el sustrato molecular del aprendizaje y la memoria a largo plazo.
- Desarrollo y maduración del SNC. Durante el desarrollo embrionario y postnatal, la actividad glutamatérgica guía la formación de sinapsis, la proliferación de espinas dendríticas y la poda sináptica competitiva. Alteraciones en esta señalización durante el neurodesarrollo se han relacionado con trastornos como el autismo y la esquizofrenia.
- Sensibilización central y dolor crónico. En el asta posterior de la médula espinal, la activación sostenida de los receptores NMDA por estímulos nociceptivos repetidos produce una forma de LTP espinal que amplifica la transmisión del dolor: este fenómeno, llamado sensibilización central, contribuye al mantenimiento del dolor crónico.
Glutamato y dolor crónico: la sensibilización central mediada por NMDA explica por qué, en síndromes de dolor crónico como la fibromialgia o la neuropatía postherpética, estímulos que normalmente no producen dolor (tacto, temperatura suave) se perciben como dolorosos (alodinia). La ketamina, antagonista del receptor NMDA, se utiliza como analgésico en estos contextos precisamente por este mecanismo.
20.7. Potenciación a largo plazo (LTP)
La potenciación a largo plazo es el mecanismo celular más estudiado del aprendizaje y la memoria. Su inducción depende de la activación del receptor NMDA; su expresión depende de cambios en los receptores AMPA de la membrana postsináptica.
Secuencia molecular de la LTP:
- La activación repetida de la sinapsis libera glutamato en cantidad suficiente para despolarizar la membrana postsináptica (vía AMPA).
- Esa despolarización retira el bloqueo por Mg²⁺ del receptor NMDA, que ya tiene glutamato y coagonista (D-serina o glicina) unidos: el canal se abre.
- Entra Ca²⁺ masivamente a través del NMDA en la espina dendrítica.
- El Ca²⁺ activa la CaMKII (calmodulina-quinasa II dependiente de Ca²⁺), que fosforila la subunidad GluA1 de los receptores AMPA.
- La fosforilación de GluA1 actúa como señal de tráfico: los receptores AMPA almacenados en vesículas intracelulares se insertan en la membrana postsináptica.
- El resultado es una mayor densidad de receptores AMPA activos en la sinapsis: la misma cantidad de glutamato produce ahora una respuesta postsináptica mayor. La sinapsis queda potenciada.
La LTP no se produce porque los receptores AMPA existentes se vuelvan «más sensibles», sino porque aumenta su número en la membrana postsináptica. Más receptores AMPA = mayor corriente despolarizante ante el mismo estímulo = sinapsis más eficaz. Este es el sustrato molecular del aprendizaje sináptico.
La LTP puede mantenerse desde segundos hasta horas o días (LTP temprana, dependiente solo de fosforilación) o semanas (LTP tardía, que requiere síntesis proteica y cambios estructurales en la espina dendrítica). Esta distinción es relevante para entender la consolidación de la memoria.
El mecanismo de la LTP es uno de los contenidos más preguntados en el MIR en el bloque de fisiología neuronal. La secuencia a memorizar es:
estimulación repetida → despolarización → retirada Mg²⁺ → apertura NMDA → entrada Ca²⁺ → CaMKII → inserción de AMPA → potenciación.
20.6. Excitotoxicidad
El glutamato, siendo el principal excitador del SNC, es también potencialmente letal para las neuronas cuando se acumula en exceso. Este fenómeno se denomina excitotoxicidad.
Excitotoxicidad: proceso de muerte neuronal causado por la sobreactivación de receptores ionotrópicos de glutamato, principalmente NMDA, que produce una entrada masiva y sostenida de Ca²⁺ en la célula, activando múltiples vías de daño celular que culminan en muerte neuronal.
La entrada masiva de Ca²⁺ no actúa por una sola vía, sino que activa en paralelo varios mecanismos de daño:
- Las calpaínas son proteasas intracelulares activadas directamente por el Ca²⁺ que degradan proteínas del citoesqueleto, de la membrana y de las mitocondrias de forma rápida y no apoptótica.
- La óxido nítrico sintetasa neuronal (nNOS) también se activa por Ca²⁺-calmodulina y produce óxido nítrico en exceso, que genera radicales libres altamente reactivos.
- Las mitocondrias sobrecargan su capacidad tampón de Ca²⁺, lo que puede abrir el poro de transición de permeabilidad mitocondrial y desencadenar la liberación de citocromo c y la activación de caspasas (vía apoptótica).
El resultado final, necrosis o apoptosis, depende de la intensidad y la velocidad de la agresión: la necrosis predomina en situaciones de daño masivo y rápido; la apoptosis, en situaciones de daño moderado o prolongado.
Excitotoxicidad en el ictus isquémico: cuando una arteria cerebral se ocluye, las neuronas del núcleo isquémico mueren rápidamente por falta de ATP y liberan su contenido intracelular, incluido el glutamato almacenado en sus terminales. El glutamato extracelular en exceso activa masivamente los receptores NMDA de las neuronas vecinas en la zona de penumbra isquémica, produciendo entrada de Ca²⁺ y muerte neuronal secundaria por excitotoxicidad. Si la glía puede recaptar el glutamato (EAAT2 funcionante, aporte de O₂ suficiente), el daño queda circunscrito al núcleo. Si no, la penumbra se incorpora al infarto.
Este mecanismo explica por qué cada minuto cuenta en el ictus y es la razón por la que se investigan antagonistas del receptor NMDA como neuroprotectores.
La excitotoxicidad no mata neuronas solo por apoptosis vía caspasas. En el núcleo isquémico, la muerte es principalmente necrótica (rápida, masiva). En la penumbra, pueden coexistir necrosis y apoptosis. Reducir la excitotoxicidad a «activación de caspasas» es una simplificación que puede generar errores en preguntas de fisiopatología.
Apoptosis, necrosis y caspasas se desarrollan con mayor profundidad en el tema de envejecimiento y muerte celular de la asignatura de Biología.
