2.1. Diferencias fundamentales entre oogénesis y espermatogénesis
Aunque ambas son formas de gametogénesis y comparten el mecanismo básico de la meiosis, la oogénesis y la espermatogénesis difieren en aspectos fundamentales que tienen consecuencias fisiológicas y clínicas importantes.
La espermatogénesis es un proceso continuo que comienza en la pubertad y produce millones de gametos por día durante toda la vida adulta. La oogénesis, en cambio, es un proceso discontinuo que comienza en la vida fetal, entra en dos arretos meióticos sucesivos que pueden durar décadas, y produce un único gameto maduro por ciclo menstrual.
La espermatogénesis es simétrica: cada espermatocito primario produce cuatro espermátidas iguales. La oogénesis es asimétrica: cada ovocito primario produce un único ovocito maduro funcional y hasta tres corpúsculos polares que no son fecundables. Esta asimetría sirve para conservar la mayor cantidad de citoplasma posible en el único gameto viable, que deberá nutrir al embrión en sus primeras divisiones.
La asimetría de las divisiones meióticas en la oogénesis no es un fallo del sistema: es una estrategia adaptativa. El ovocito necesita acumular reservas de ARN, proteínas y orgánulos que sostendrán el desarrollo embrionario hasta que el genoma del cigoto comience a expresarse (activación del genoma embrionario, hacia el estadio de 8 células en humanos).
| Espermatogénesis | Oogénesis | |
|---|---|---|
| Inicio | Pubertad | Vida fetal (semana 8-9) |
| Ritmo | Continuo (millones/día) | Discontinuo (1/ciclo) |
| Arresto meiótico | No hay | Dos arretos (profase I y metafase II) |
| Productos por gameto primario | 4 gametos funcionales | 1 gameto funcional + hasta 3 corpúsculos polares |
| Localización | Túbulos seminíferos | Folículos ováricos |
| Fin | No cesa (toda la vida adulta) | Menopausia (~50 años) |
2.2. Fases de la oogénesis
2.2.1. Oogonias y primer arresto meiótico
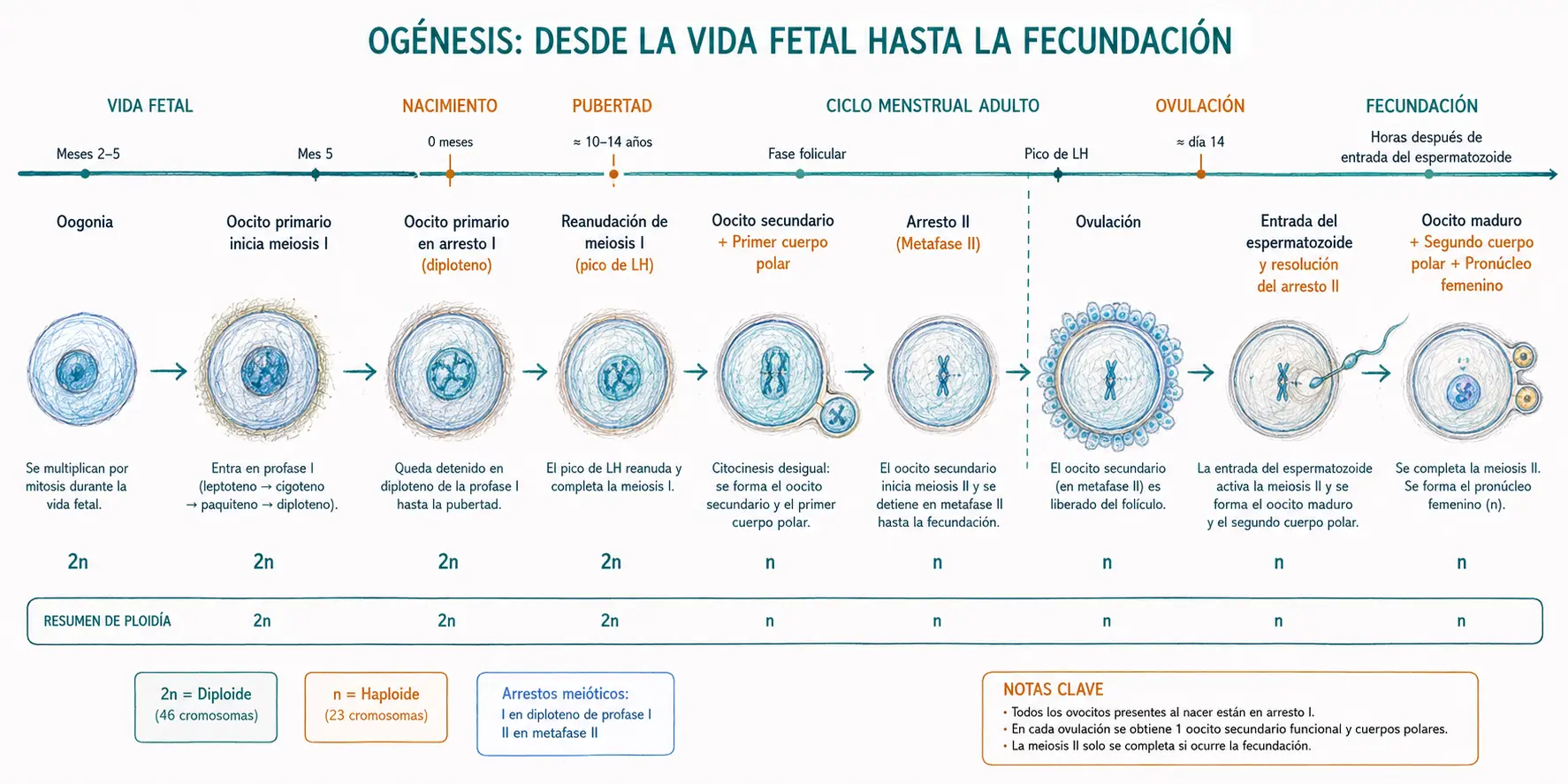
Las células germinales femeninas llegan al esbozo gonadal durante el desarrollo embrionario y se denominan oogonias. En la gónada femenina, las oogonias proliferan activamente por mitosis durante los meses 2 a 5 de la vida fetal, alcanzando un máximo de aproximadamente 6-7 millones de células en torno a la semana 20.
A partir del quinto mes de vida fetal, las oogonias comienzan a entrar en meiosis I, transformándose en ovocitos primarios (2n, 4c). Sin embargo, la meiosis I no se completa: los ovocitos primarios se detienen en el diploteno de la profase I, un estadio en el que los cromosomas homólogos están ya emparejados y se ha producido el sobrecruzamiento, pero aún no ha ocurrido la separación.
El primer arresto meiótico es la detención del ovocito primario en el estadio de diploteno de la profase I. Se mantiene gracias a la acción de factores inhibidores producidos por las células foliculares que rodean al ovocito, en particular el AMPc. Este arresto puede prolongarse desde los meses fetales hasta la pubertad, y en cada ovocito se mantiene hasta el pico preovulatorio de LH.
Al mismo tiempo que los ovocitos primarios entran en arresto meiótico, cada uno queda rodeado por una capa de células epiteliales planas, las células foliculares, formando las unidades básicas denominadas folículos primordiales. Este proceso se completa antes del nacimiento. Al nacer, la niña tiene entre 1 y 2 millones de folículos primordiales; para la pubertad, solo quedan unos 300.000-400.000, el resto ha sufrido atresia.
La reserva ovárica queda fijada definitivamente antes del nacimiento. No hay formación de nuevos ovocitos durante la vida postnatal en condiciones fisiológicas. Esta limitación explica el envejecimiento ovárico y la menopausia: cuando la reserva de folículos se agota o cae por debajo de un umbral crítico, el ciclo menstrual cesa.
2.2.2. Desarrollo folicular
El desarrollo folicular comprende una serie de transformaciones estructurales en el folículo que acompañan a la maduración del ovocito.
- El folículo primordial consiste en un ovocito primario rodeado de una sola capa de células foliculares planas. La mayoría de los folículos primordiales permanecen en este estadio durante años. Periódicamente, cohortes de folículos primordiales son reclutadas para iniciar el desarrollo; de ellas, la inmensa mayoría sufrirá atresia.
- En el folículo primario, las células foliculares adquieren morfología cúbica y proliferan formando varias capas, denominadas ahora células de la granulosa. El ovocito comienza a crecer y secreta la zona pelúcida, una capa de glucoproteínas que lo rodea y que más tarde será el sustrato para la unión y penetración del espermatozoide.
- En los folículos secundarios, las células de la granulosa continúan proliferando y aparece el estroma folicular, que diferencia una capa de células estromales adyacentes al folículo: la teca interna (altamente vascularizada, productora de andrógenos bajo estímulo de LH) y la teca externa (fibrosa, de sostén).
- El folículo de mayor tamaño, el folículo de Graaf (o folículo maduro), se caracteriza por la presencia del antro folicular, una cavidad llena de líquido folicular que se forma por la secreción de las células de la granulosa. El ovocito queda excéntrico, suspendido en una colina de células de granulosa denominada cúmulo oóforo (u ooforus). El líquido folicular contiene estrógenos y factores de crecimiento que regulan la función del propio folículo.
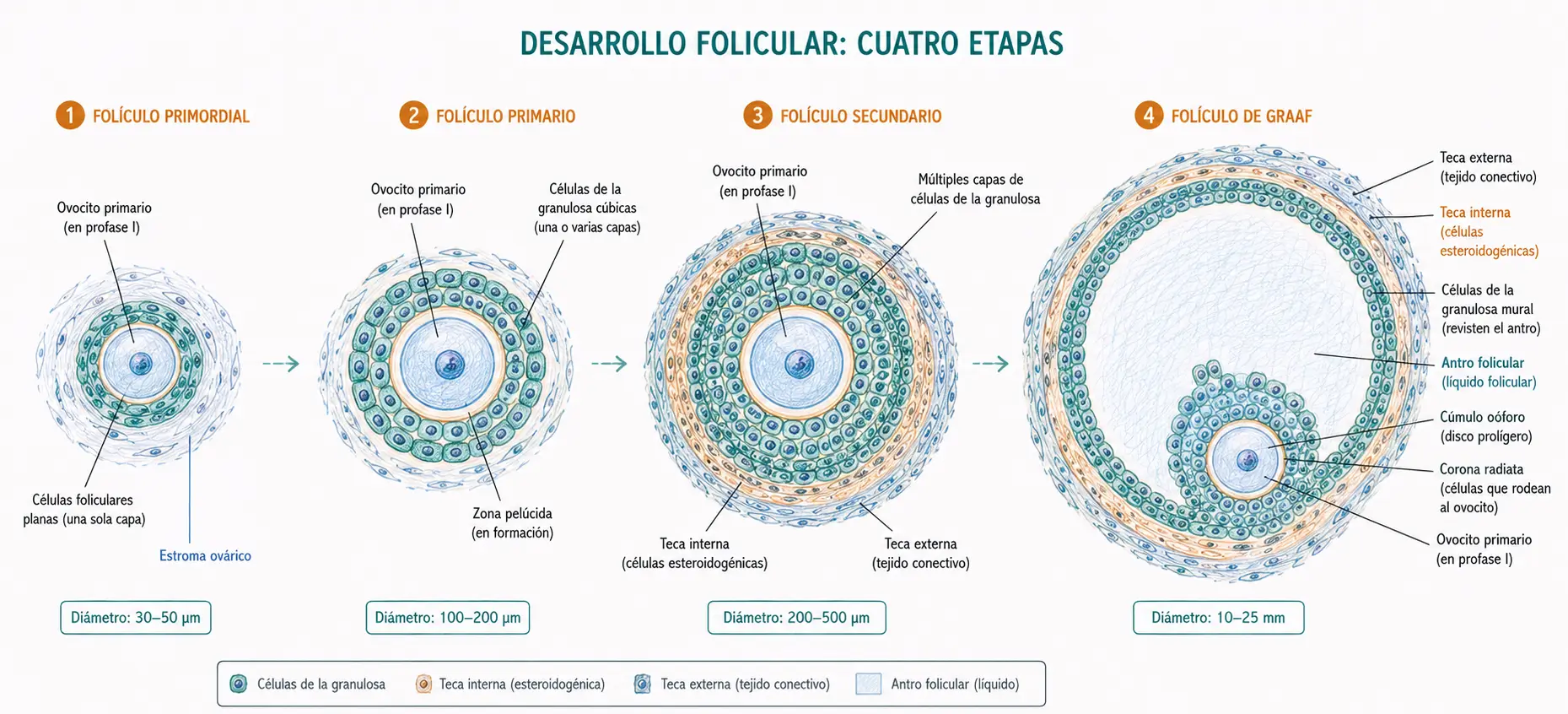
La zona pelúcida está formada principalmente por tres glucoproteínas: ZP1, ZP2 y ZP3. La ZP3 actúa como receptor del espermatozoide e inicia la reacción acrosómica. Tras la fecundación, la zona pelúcida se endurece (reacción de zona) para impedir la polispermia.
En cada ciclo menstrual, la cohorte de folículos reclutada habitualmente origina un único folículo dominante que supera la atresia. Los demás folículos de la cohorte degeneran. El folículo dominante es el que ovulará. Los embarazos gemelares dicigóticos ocurren cuando dos folículos ovulan en el mismo ciclo.
2.3. Ovulación y resolución del primer arresto
El pico preovulatorio de LH desencadenado a mitad del ciclo menstrual es el estímulo que resuelve el primer arresto meiótico. Bajo el efecto de la LH, el ovocito primario reanuda la meiosis I, que se completa en pocas horas produciendo dos células de tamaño muy desigual: el ovocito secundario (n, 2c), que recibe casi todo el citoplasma, y el primer corpúsculo polar (n, 2c), una célula pequeña que no tiene relevancia funcional y que posteriormente puede o no dividirse.
Inmediatamente después de completar la meiosis I, el ovocito secundario inicia la meiosis II pero se detiene de nuevo en la metafase II: es el segundo arresto meiótico. En este estadio se produce la ovulación: el folículo de Graaf se rompe y el ovocito secundario, rodeado del cúmulo oóforo, es expulsado del ovario.
Lo que se ovula no es un ovocito maduro, sino un ovocito secundario arrestado en metafase II. La maduración final del ovocito, con la extrusión del segundo corpúsculo polar, solo ocurre si hay fecundación. Si no hay fecundación, el ovocito secundario degenera sin completar la meiosis II.
Tras la ovulación, el ovocito secundario es captado por las fimbrias de la trompa de Falopio y comienza su desplazamiento hacia la cavidad uterina. La fecundación, si ocurre, tiene lugar en el tercio externo de la trompa (ampolla tubárica), habitualmente en las 12-24 horas siguientes a la ovulación.
2.4. Resolución del segundo arresto: la fecundación
La penetración del espermatozoide en el ovocito secundario es el estímulo que desencadena la resolución del segundo arresto meiótico. La entrada del espermatozoide activa una cascada de señalización intracelular mediada por calcio que permite que la meiosis II se complete: el ovocito secundario se divide asimétricamente en el ovocito maduro (n, 1c) y el segundo corpúsculo polar (n, 1c).
Tras la extrusión del segundo corpúsculo polar, el núcleo del ovocito maduro se denomina pronúcleo femenino. El núcleo del espermatozoide se descondensa y forma el pronúcleo masculino. La fusión de ambos pronúcleos constituye la anfimixis, que restablece la ploídía diploide (2n = 46) y marca el inicio de la vida del nuevo individuo.
La fecundación es la fusión del gameto masculino (espermatozoide) con el gameto femenino (ovocito secundario) que restablece la ploídía diploide y activa el desarrollo embrionario. Sus resultados son:
- Restauración de 2n = 46 cromosomas.
- Determinación del sexo genético del nuevo individuo.
- Inicio de las divisiones de segmentación del cigoto.
El proceso completo de fecundación, la primera semana de desarrollo y los mecanismos de prevención de la polispermia se desarrollan en el Tema 8 · Fecundación.
2.5. Regulación hormonal del ciclo ovárico
La oogénesis en la mujer adulta está encuadrada en el ciclo menstrual, un ciclo de aproximadamente 28 días regulado por el eje hipotálamo-hipófisis-ovario y sincronizado con los cambios del endometrio uterino.
La regulación detallada del ciclo menstrual, incluyendo las fases folicular, ovulatoria y lútea, los cambios endometriales y la fisiología de los estrógenos y la progesterona, se estudia en Fisiología (→ Fisiología: regulación del ciclo reproductor femenino) y se amplía en Ginecología y Obstetricia.
En el contexto de la gametogénesis interesa destacar los momentos de la regulación hormonal ovárica:
- Durante la fase folicular, la FSH hipofisaria estimula el crecimiento folicular y la producción de estrógenos por las células de la granulosa. Los estrógenos ejercen retroalimentación positiva sobre la hipófisis cuando alcanzan niveles elevados, lo que desencadena el pico de LH.
- El pico de LH a mitad del ciclo (día ~14 en un ciclo de 28 días) es el desencadenante directo de la ovulación y de la resolución del primer arresto meiótico del ovocito primario.
- Tras la ovulación, las células de la granulosa y la teca interna se luteinizan bajo el efecto de la LH, formando el cuerpo lúteo, que secreta progesterona.
- La progesterona prepara el endometrio para la implantación. Si no hay implantación, el cuerpo lúteo degenera al cabo de 14 días, los niveles de progesterona y estrógenos caen, y se produce la menstruación.
La estimulación ovárica controlada en reproducción asistida se basa en la administración exógena de FSH para reclutar y madurar simultáneamente múltiples folículos (en condiciones naturales solo uno llega a ovular). Cuando los folículos alcanzan el tamaño adecuado, se administra hCG (que mimetiza el pico de LH) para desencadenar la maduración final y programar la punción folicular 36 horas después, justo antes de que ocurra la ovulación espontánea.
2.6. Atresia folicular y envejecimiento ovárico
La gran mayoría de los folículos reclutados en cada ciclo no llegan a ovular: sufren atresia, un proceso de apoptosis programada que afecta tanto al ovocito como a las células foliculares. La atresia es el destino fisiológico normal de más del 99,9% de los folículos.
La reserva folicular disminuye de forma continua e irreversible a lo largo de la vida: de los 6-7 millones de células germinales en el pico fetal a 1-2 millones al nacer, unos 300.000-400.000 en la pubertad y aproximadamente 1.000 en torno a la menopausia. Cuando la reserva cae por debajo de este umbral crítico, el ciclo menstrual cesa: es la menopausia, que ocurre en promedio alrededor de los 50 años.
La reserva ovárica es un concepto clínico que refleja la cantidad y calidad de los ovocitos restantes. Se evalúa mediante la determinación de la hormona antimülleriana (AMH) sérica y el recuento de folículos antrales por ecografía. Una reserva ovárica disminuida se asocia a menor respuesta a la estimulación ovárica en reproducción asistida y a menor fertilidad espontánea. La edad materna es el principal factor de deterioro de la reserva y la calidad ovocitaria.
El envejecimiento ovárico no solo implica una reducción cuantitativa de los folículos: también conlleva una mayor tasa de errores en la segregación cromosómica durante la meiosis I, lo que explica el aumento de aneuploidías embrionarias (como la trisomía 21) con la edad materna. Este deterioro cualitativo es el principal responsable de la disminución de la fertilidad y el aumento de abortos espontáneos en mujeres mayores de 35-38 años.
