9.1. Las divisiones de segmentación
Tras la fecundación, el cigoto inicia una serie de divisiones mitóticas denominadas divisiones de segmentación o clivaje. Estas divisiones tienen una característica fundamental que las distingue de las divisiones mitóticas del organismo adulto: no van seguidas de una fase de crecimiento celular. Cada división produce células más pequeñas, llamadas blastómeros, de modo que el embrión se divide en un número creciente de células sin aumentar su volumen total.
Durante la segmentación, el embrión no crece. El objetivo no es aumentar la masa celular sino incrementar el número de células y reducir su tamaño hasta aproximarlo al de las células somáticas normales. El embrión permanece contenido dentro de la zona pelúcida y utiliza como fuente de energía las reservas acumuladas en el citoplasma del ovocito.
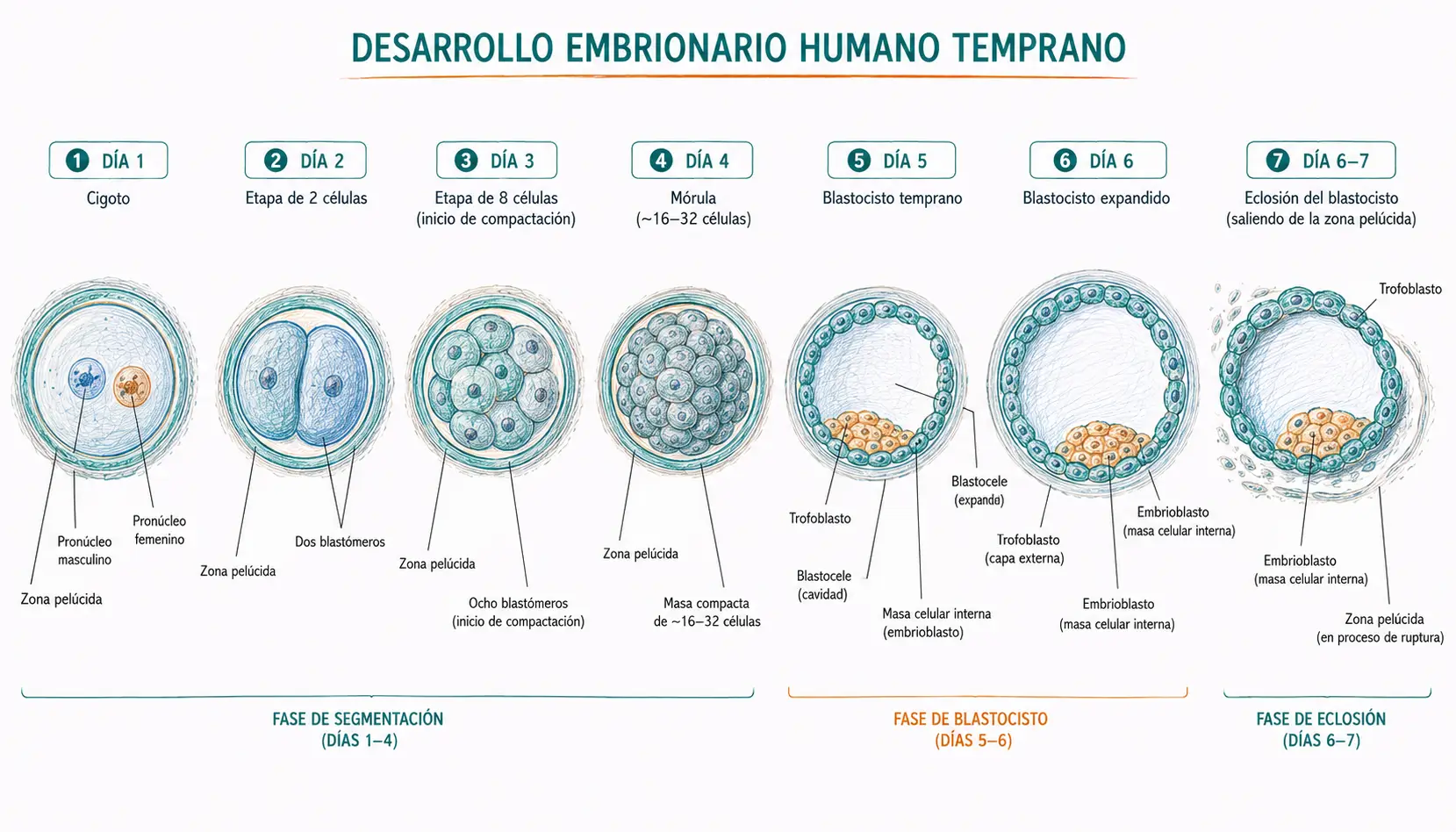
Las primeras divisiones son lentas (cada división dura unas 12-24 horas) y asincrónicas: no todos los blastómeros se dividen al mismo tiempo, por lo que el embrión puede pasar transitoriamente por estadios de 3, 5 o 7 células. Durante las primeras divisiones, todos los blastómeros son totipotentes: cada uno tiene capacidad de originar un individuo completo si se separa de los demás. Esta totipotencia se pierde progresivamente a medida que avanza la segmentación.
La totipotencia de los blastómeros tempranos es la base de la formación de gemelos monocigóticos: si el embrión se divide espontáneamente en los primeros días, cada fragmento puede originar un individuo genéticamente idéntico. También es el fundamento del diagnóstico genético preimplantacional (DGP), en el que se extrae uno o dos blastómeros del embrión en estadio de 6-8 células para analizarlos genéticamente sin comprometer el desarrollo del embrión restante.
Las primeras divisiones producen los estadios de 2, 4 y 8 células. En torno al estadio de 8 células se produce un evento crucial: la activación del genoma embrionario (o transición materno-cigótica). Hasta ese momento, el desarrollo había sido dirigido por los ARNm y proteínas maternos acumulados en el ovocito durante la oogénesis. A partir de las 8 células, el embrión comienza a transcribir su propio genoma y pasa a controlar su propio desarrollo.
9.2. Compactación y formación de la mórula
Hacia el estadio de 8-16 células, los blastómeros experimentan la compactación: las células se aplanan entre sí, maximizando el contacto intercelular. La compactación está mediada por la E-caderina, una proteína de adhesión celular calcio-dependiente expresada en la membrana de los blastómeros. La compactación es el primer evento morfogenético del desarrollo embrionario: a partir de ella, las células empiezan a diferenciarse según su posición.
Las células externas de la mórula compactada están en contacto con el exterior y desarrollarán características epiteliales: establecerán uniones oclusivas entre ellas y polarizarán su membrana. Las células internas, completamente rodeadas por las externas, recibirán señales diferentes y seguirán un destino distinto.
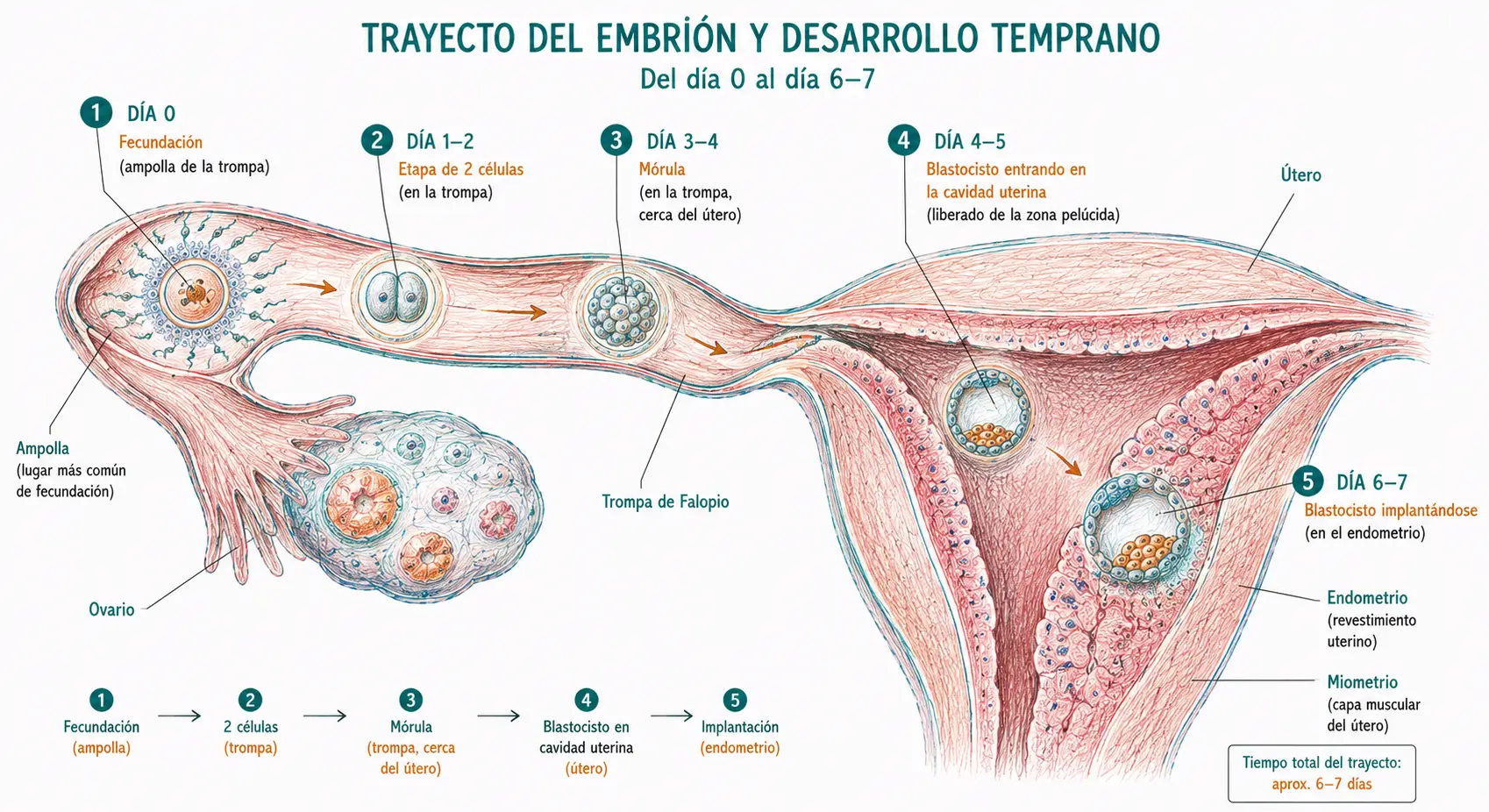
La mórula (del latín mora, mora, por su aspecto) es el estadio embrionario de 16-32 células, caracterizado por ser una masa celular compacta sin cavidad interna. Se forma alrededor del día 3-4 del desarrollo y se encuentra todavía dentro de la zona pelúcida, en tránsito por la trompa de Falopio hacia el útero.
9.3. Cavitación y formación del blastocisto
A partir del día 4, las células externas de la mórula (con sus uniones oclusivas establecidas) comienzan a bombear activamente iones sodio hacia el interior de la masa celular. El agua entra por ósmosis siguiendo el gradiente iónico y se acumula en pequeñas cavidades entre las células internas. Estas cavidades confluyen progresivamente hasta formar una única cavidad central: el blastocele.
La formación del blastocele marca el inicio del estadio de blastocisto. En este momento se establecen con claridad los dos primeros linajes celulares del desarrollo:
- El trofoblasto es la capa externa de células que rodea el blastocele. Deriva de las células externas de la mórula. Dará lugar a la placenta, el corion y las demás membranas extraembrionarias. El trofoblasto nunca formará parte del embrión propiamente dicho.
- La masa celular interna (MCI) o embrioblasto es un grupo de células que se acumulan en uno de los polos del blastocisto, el polo embrionario. Deriva de las células internas de la mórula. De ella derivarán el embrión y todos los tejidos fetales, incluyendo el amnios y el saco vitelino.
La separación entre trofoblasto y embrioblasto es la primera decisión de destino celular del desarrollo humano. Las células del trofoblasto nunca podrán convertirse en células del embrión, y viceversa. Esta restricción está regulada por factores de transcripción específicos: OCT4 y NANOG en el embrioblasto, CDX2 en el trofoblasto.
Las células del embrioblasto son pluripotentes: pueden originar cualquier tipo celular del organismo adulto, pero no pueden originar placenta. Las células del trofoblasto son multipotentes con restricción de linaje: solo pueden originar tipos celulares placentarios. Las células madre embrionarias (ES cells) de uso en investigación se derivan precisamente del embrioblasto del blastocisto humano, de ahí su pluripotencia.
9.4. Eclosión del blastocisto
Mientras el blastocisto crece y el blastocele se expande, la zona pelúcida se adelgaza progresivamente. Hacia el día 5-6, el blastocisto produce enzimas (entre ellas la estapsilisina, o estrepsilisina blastocística) que digieren localmente la zona pelúcida, y la presión interna del blastocele termina de romperla. El blastocisto emerge al exterior en un proceso denominado eclosión.
La eclosión es un requisito imprescindible para la implantación: mientras el blastocisto esté encerrado en la zona pelúcida, el trofoblasto no puede entrar en contacto con el epitelio endometrial.
En los ciclos de fertilización in vitro (FIV), algunos blastocistos no eclosionan espontáneamente en el laboratorio, lo que puede reducir las tasas de implantación. La eclosión asistida (assisted hatching) es una técnica en la que el embriólogo practica una pequeña apertura en la zona pelúcida mediante láser, ácido de Tyrode o aguja mecánica para facilitar la eclosión. Su beneficio clínico es objeto de debate, pero se utiliza habitualmente en embriones con zona pelúcida gruesa o en pacientes con fallos repetidos de implantación.
9.5. El endometrio y la ventana de implantación
Para que la implantación pueda producirse, no solo el blastocisto debe estar preparado: el endometrio debe encontrarse en un estado receptivo preciso. Esta receptividad endometrial se denomina ventana de implantación y ocurre entre los días 20 y 24 del ciclo menstrual (días 6-10 postovulatorios).
Durante la fase folicular del ciclo, los estrógenos producidos por el folículo en crecimiento estimulan la proliferación del epitelio endometrial. Tras la ovulación, la progesterona del cuerpo lúteo induce la diferenciación secretora del endometrio: las glándulas endometriales se hacen tortuosas y secretoras, el estroma se edematiza y las arterias espirales se desarrollan. Todo ello prepara un lecho vascular y nutritivo para la implantación.
Durante la ventana de implantación, el epitelio endometrial expresa en su superficie una serie de moléculas de adhesión, entre ellas las pinópodas (proyecciones apicales del epitelio), la osteopontina y las integrinas, que facilitan el anclaje inicial del trofoblasto.
La ventana de implantación es estrecha y precisa. Si el blastocisto llega demasiado pronto (endometrio en fase no receptiva) o demasiado tarde (endometrio ya fuera de la ventana), la implantación no ocurre y el embrión se pierde. La sincronía entre el desarrollo embrionario y la preparación endometrial es uno de los principales determinantes del éxito reproductivo.
9.6. Inicio de la implantación y diferenciación del trofoblasto
Hacia el día 6-7 del desarrollo, el blastocisto eclosionado toma contacto con el epitelio endometrial en la región del polo embrionario (donde está el embrioblasto). La implantación se inicia con tres pasos secuenciales: aposición, adhesión e invasión.
- En la aposición, el blastocisto se aproxima y orienta sobre el epitelio endometrial, con el polo embrionario dirigido hacia el epitelio.
- En la adhesión, el trofoblasto establece uniones firmes con las células epiteliales endometriales mediante integrinas y otras moléculas de adhesión.
- En la invasión, las células del trofoblasto comienzan a penetrar activamente entre las células epiteliales endometriales.
Al final de la primera semana, el blastocisto ha comenzado a introducirse en el endometrio y el trofoblasto se ha diferenciado en dos capas:
- El citotrofoblasto: capa interna de células individuales con actividad mitótica.
- El sincitiotrofoblasto: capa externa formada por la fusión de células del citotrofoblasto, que forma un sincitio multinucleado con potente capacidad invasiva.
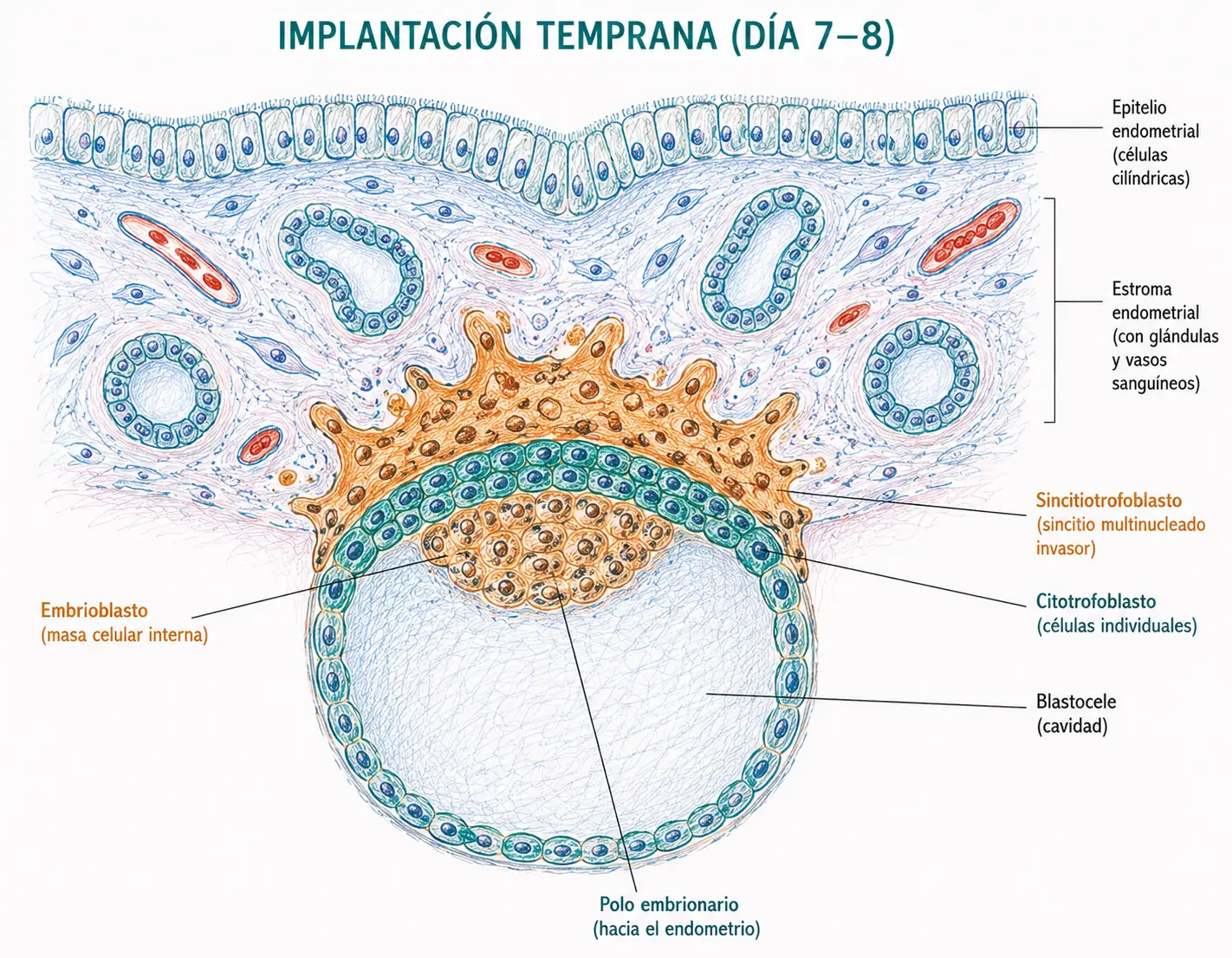
El sincitiotrofoblasto es la célula invasiva por excelencia del embrión temprano: secreta enzimas proteolíticas que digieren la matriz extracelular y las uniones entre las células endometriales, abriendo camino para la implantación. También es la primera fuente de hCG (gonadotropina coriónica humana), la hormona que mantiene el cuerpo lúteo activo y que es la base de los tests de embarazo.
La hCG (gonadotropina coriónica humana) es producida por el sincitiotrofoblasto desde el inicio de la implantación, aproximadamente a partir del día 8-10 del desarrollo. Sus niveles en sangre y orina se duplican aproximadamente cada 48 horas durante las primeras semanas del embarazo. Los tests de embarazo comerciales detectan hCG en orina a partir de niveles de 20-25 mUI/mL, lo que corresponde aproximadamente al día 10-14 tras la ovulación. La determinación cuantitativa de hCG en sangre permite además el seguimiento de la evolución del embarazo en sus fases iniciales.
El proceso completo de implantación, incluyendo la reacción decidual del endometrio y el desarrollo de las vellosidades coriónicas, se desarrolla durante la segunda semana y se estudia en el Tema 10 · Segunda semana de desarrollo.
