21.1. Introducción
Hacia la cuarta semana de desarrollo, en la porción distal del intestino anterior, aparece una evaginación que constituye el punto de partida de todo el aparato respiratorio: el primordio o divertículo respiratorio.
El intestino anterior es la misma estructura que da origen al esófago, al estómago y a los primeros derivados hepatopancreáticos (ver Tema 22 · Aparato digestivo). La proximidad topográfica entre esófago y vía respiratoria en este estadio explica que ambos compartan mecanismo de separación.
21.2. Formación del primordio respiratorio
En la cara ventral de la faringe primitiva aparece el surco laringotraqueal, que se profundiza progresivamente hasta formar el divertículo laringotraqueal, el cual crece en sentido ventrocaudal separándose del intestino anterior.
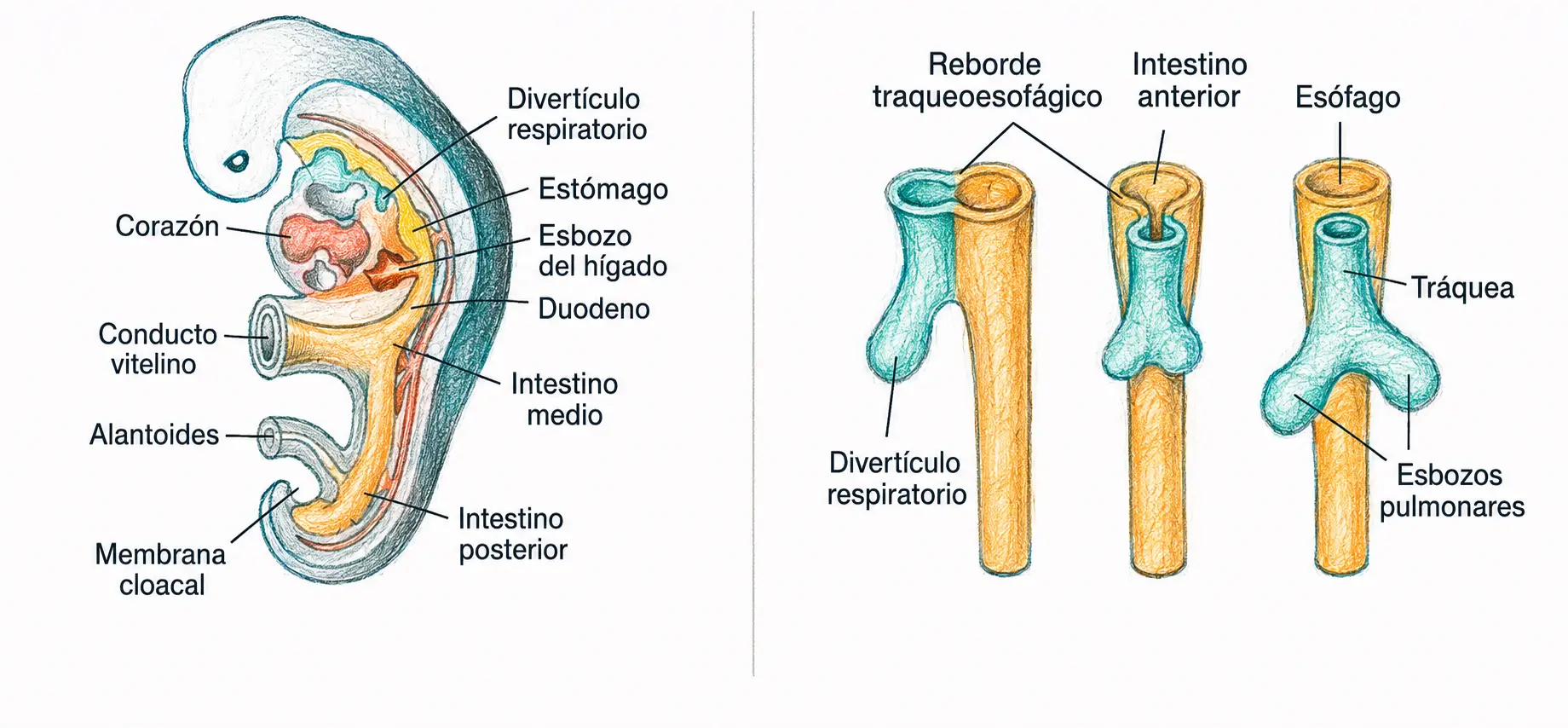
Tabique traqueoesofágico: proceso de pinzamiento que separa definitivamente el esófago (dorsal) del tubo laringotraqueal (ventral), ambos originados en la misma evaginación inicial del intestino anterior.
Un fallo parcial o completo del tabique traqueoesofágico deja una comunicación anómala entre tráquea y esófago: la fístula traqueoesofágica, con frecuencia asociada a atresia esofágica. Es una de las malformaciones congénitas del aparato digestivo/respiratorio con mayor relevancia clínica neonatal.
21.3. Desarrollo de la laringe
El esqueleto cartilaginoso de la laringe no se forma a partir de tejido respiratorio propio, sino del mesénquima de los arcos faríngeos 4º y 6º, ya estudiados en el tema 18.
El mesénquima de estos dos arcos prolifera y forma las protuberancias aritenoideas, que originan la epiglotis y, en su interior, los cartílagos laríngeos definitivos (tiroides, cricoides, aritenoides). La epiglotis, en concreto, se desarrolla a partir de la porción caudal de la eminencia hipobranquial (ver tema 19, apartado de formación de la lengua, misma estructura embrionaria).
Durante su desarrollo, la luz de la laringe se cierra transitoriamente por proliferación del propio epitelio de revestimiento. Hacia el tercer mes, esta luz se reabre y se forman dos divertículos laríngeos laterales que darán origen a las cuerdas vocales.
21.4. Desarrollo de tráquea, bronquios y pulmones
A partir del tubo laringotraqueal se desarrollan, en dirección caudal, la tráquea, los bronquios y los pulmones. Como en el resto del aparato respiratorio, el epitelio de revestimiento interno es de origen endodérmico, mientras que los componentes cartilaginosos, musculares y conectivos derivan del mesénquima esplácnico circundante (esplacnopleura).
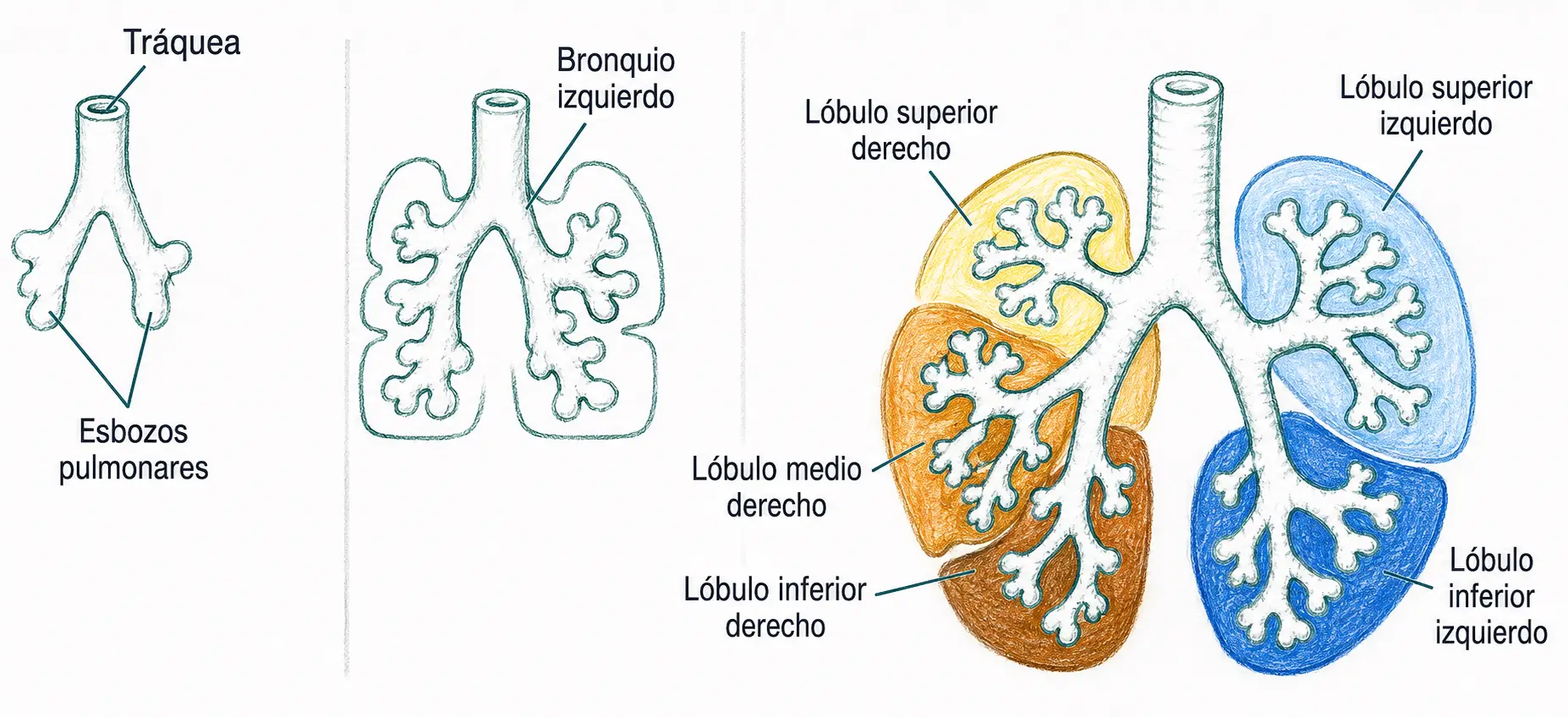
En el tubo laringotraqueal aparecen, hacia la quinta semana, dos yemas pulmonares primarias. La yema derecha se subdivide en tres yemas secundarias y la izquierda en dos, reflejo directo de la asimetría de tres lóbulos (derecho) y dos lóbulos (izquierdo) que tendrá el pulmón adulto.
Cada yema secundaria continúa ramificándose hasta formar diez bronquios segmentarios o terciarios por pulmón. La secuencia completa de ramificación es:
Tráquea → bronquio principal → bronquios lobares → bronquios segmentarios (10 por pulmón) → bronquiolo → bronquiolo terminal → alveolo.
El epitelio de todo el árbol respiratorio, incluidos los alveolos, es de origen endodérmico, no ectodérmico. El mesénquima que rodea a los bronquiolos terminales se reorganiza para formar los propios alveolos, revestidos por dos tipos de neumocitos: tipo I (intercambio gaseoso) y tipo II (secreción de surfactante).
El mesénquima que rodea a los bronquiolos terminales no se limita a acompañar pasivamente el crecimiento del árbol bronquial, se reorganiza activamente para formar el propio parénquima pulmonar. En este proceso de reorganización aparecen las cúpulas celulares que constituirán los alveolos.
21.5. Maduración pulmonar
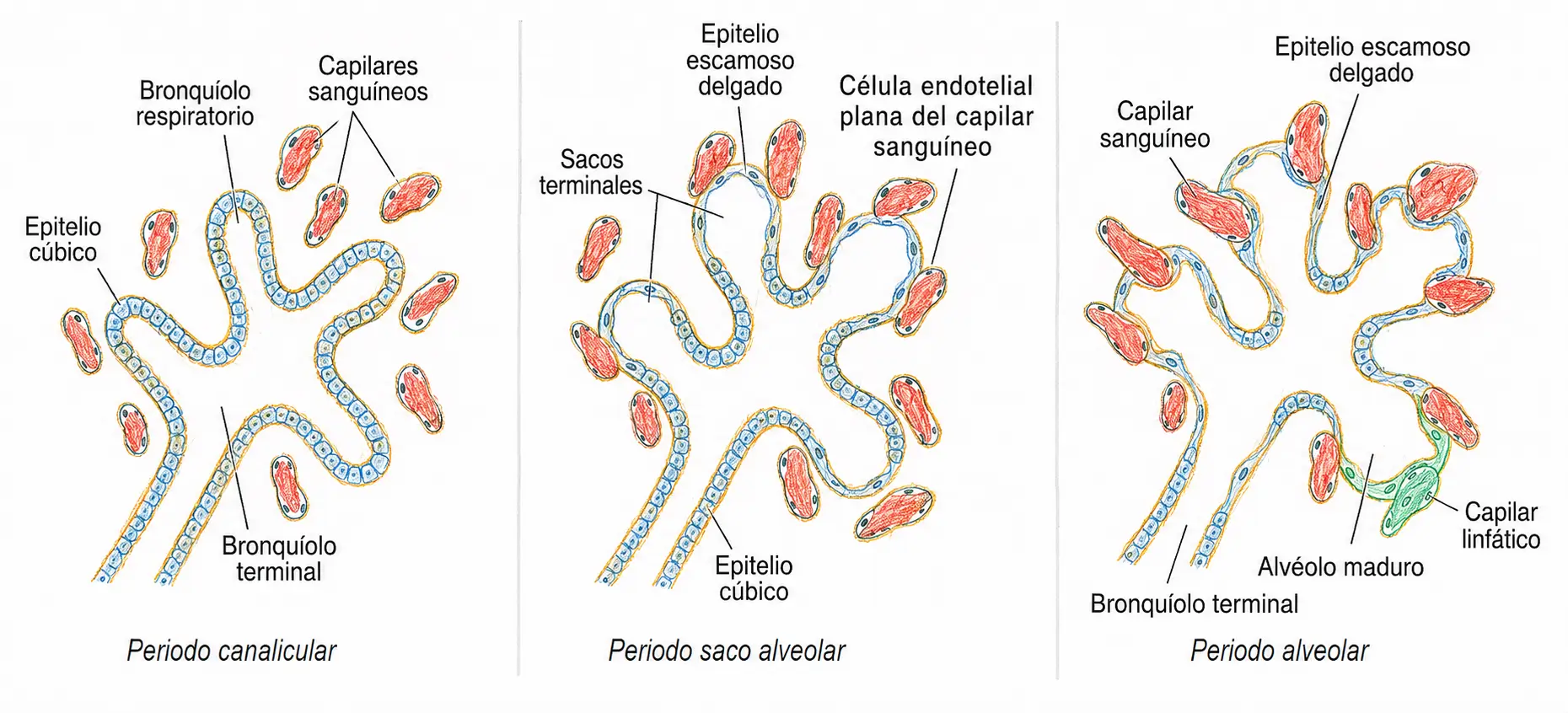
Una vez formado el árbol bronquial completo, el pulmón todavía no es funcional. La maduración histológica que lo hace apto para el intercambio gaseoso atraviesa cuatro periodos sucesivos, cada uno definido por un cambio estructural concreto en el epitelio y en la vascularización del tejido pulmonar.
Durante el periodo pseudoglandular, que se extiende hasta aproximadamente el cuarto mes, el sistema de conducción de aire queda establecido en su totalidad: desde los bronquios hasta los bronquiolos, todo el árbol respiratorio se recubre de un epitelio cúbico grueso.
Este revestimiento da al tejido un aspecto histológico similar al de una glándula exocrina, de ahí el nombre del periodo. En esta fase no existe ninguna estructura capaz de realizar intercambio gaseoso: un feto que naciera en este periodo no podría respirar bajo ningún concepto, con independencia del soporte médico disponible.
El periodo pseudoglandular no prepara al pulmón para respirar, prepara únicamente la tubería por la que circulará el aire. Todavía falta vascularizar el tejido y adelgazar el epitelio, que es lo que ocurre en los dos periodos siguientes.
Entre el cuarto y el sexto mes tiene lugar el periodo canalicular. En esta fase aumenta de forma notable el calibre de bronquios y bronquiolos, y por primera vez el tejido pulmonar se vasculariza de manera significativa. Los capilares sanguíneos se aproximan al epitelio respiratorio. Cada bronquiolo terminal se divide en dos o más bronquiolos respiratorios, que a su vez forman entre tres y seis conductos alveolares. El nombre del periodo hace referencia a la formación de canales o conductos progresivamente más finos.
A partir de la semana 30 comienza el periodo de saco alveolar, el punto de inflexión real en cuanto a viabilidad. En los conductos alveolares ya formados proliferan los propios alveolos, y es en este momento cuando arranca la síntesis y secreción de surfactante. Es también en este periodo cuando se distinguen con claridad los dos tipos de neumocitos:
- Tipo I, aplanados y especializados en el intercambio gaseoso.
- Tipo II, cuboidales y responsables de secretar el surfactante que evita el colapso alveolar.
El periodo de saco alveolar marca el límite práctico de viabilidad extrauterina, no por casualidad sino porque es el primer momento del desarrollo en el que existe surfactante disponible. Sin surfactante, la tensión superficial dentro del alveolo lo hace colapsar en cada espiración: es la base fisiopatológica del síndrome de dificultad respiratoria del recién nacido, la principal causa de morbimortalidad respiratoria en grandes prematuros.
Finalmente, el periodo alveolar se extiende desde el octavo mes hasta bien entrada la infancia, mucho más allá del nacimiento. En esta fase el epitelio alveolar se aplana todavía más, lo que aumenta de forma considerable la superficie disponible para el intercambio gaseoso, y se produce un incremento masivo en el número absoluto de alveolos.
Un recién nacido a término no completa su dotación definitiva de alveolos en el parto: ese proceso continúa activamente durante los primeros años de vida.
| Periodo | Edad gestacional | Hallazgo clave | Relevancia clínica |
|---|---|---|---|
| Pseudoglandular | Hasta el 4º mes aprox. | Se establece el sistema de conducción de aire; epitelio cúbico da aspecto glandular | Sin capacidad respiratoria: inviable si nace en este periodo |
| Canalicular | 4º a 6º mes | Aumenta el calibre bronquial, se vasculariza el tejido; cada bronquiolo terminal forma 3-6 conductos alveolares | Todavía muy limitada capacidad de intercambio gaseoso |
| Saco alveolar | Desde la semana 30 | Proliferación de alveolos, inicio de síntesis de surfactante, se diferencian neumocitos I y II | Límite práctico de viabilidad extrauterina |
| Alveolar | 8º mes hasta la infancia | El epitelio se aplana, aumenta mucho el número de alveolos | Maduración funcional plena, continúa tras el nacimiento |
Neumocito tipo I: aplanado, mayoritario en superficie, especializado en el intercambio gaseoso, sin capacidad de división.
Neumocito tipo II: cuboidal, minoritario en superficie pero más numeroso en número absoluto, secreta surfactante y actúa además como célula madre capaz de regenerar el epitelio alveolar dañado (incluido el propio tipo I).
21.6. Formación de las pleuras y el diafragma
Las yemas pulmonares, al crecer en dirección caudal y lateral, se introducen en la cavidad celómica y arrastran consigo la esplacnopleura del tubo digestivo, mientras la somatopleura recubre la pared del embrión.
De esta relación derivan las dos hojas pleurales:
- La pleura visceral, de la esplacnopleura.
- La pleura parietal, de la somatopleura.
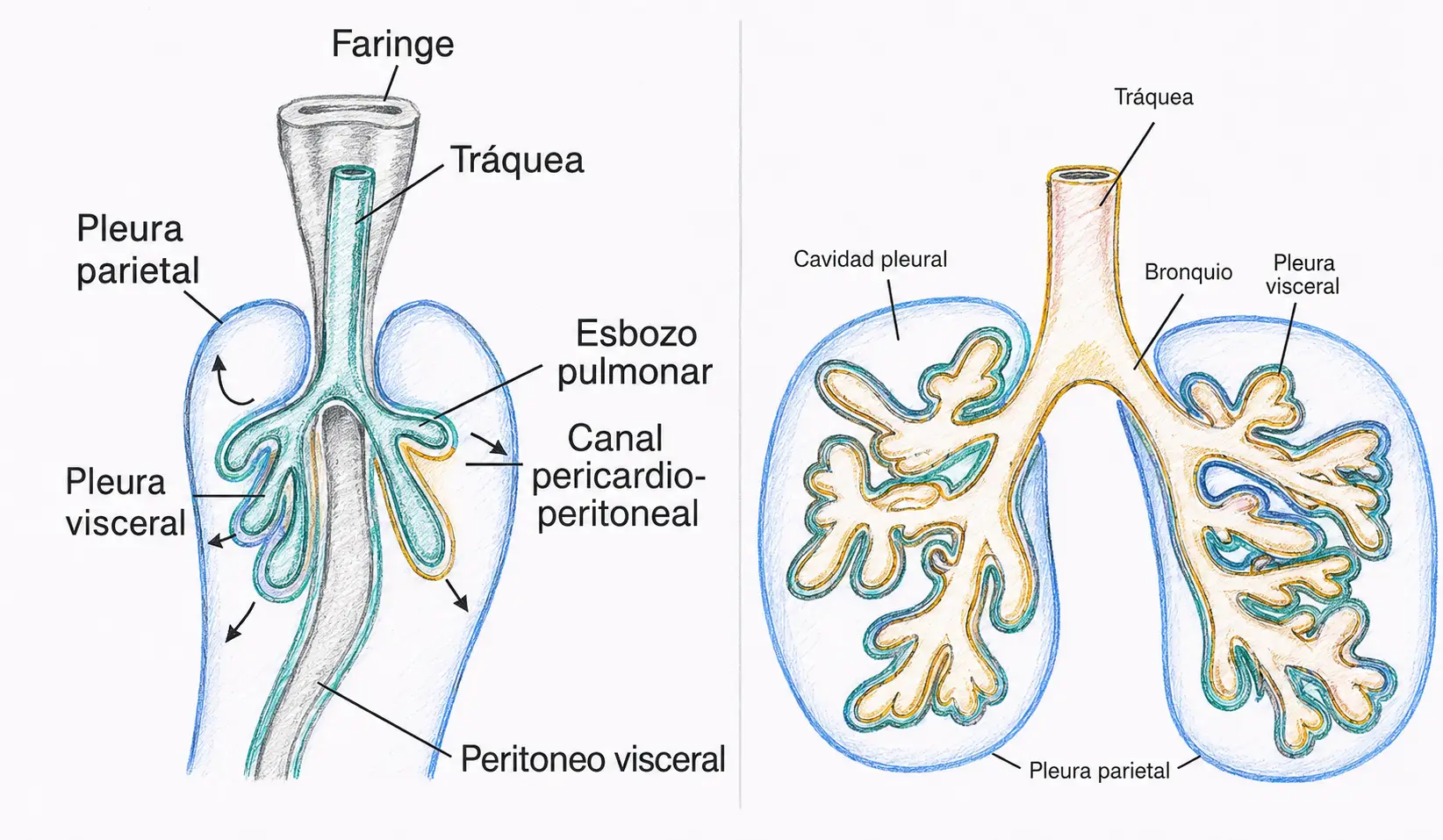
Antes de que exista un diafragma como estructura única, el corazón en desarrollo está unido a las estructuras vecinas por unos tabiques transitorios llamados mesocardios.
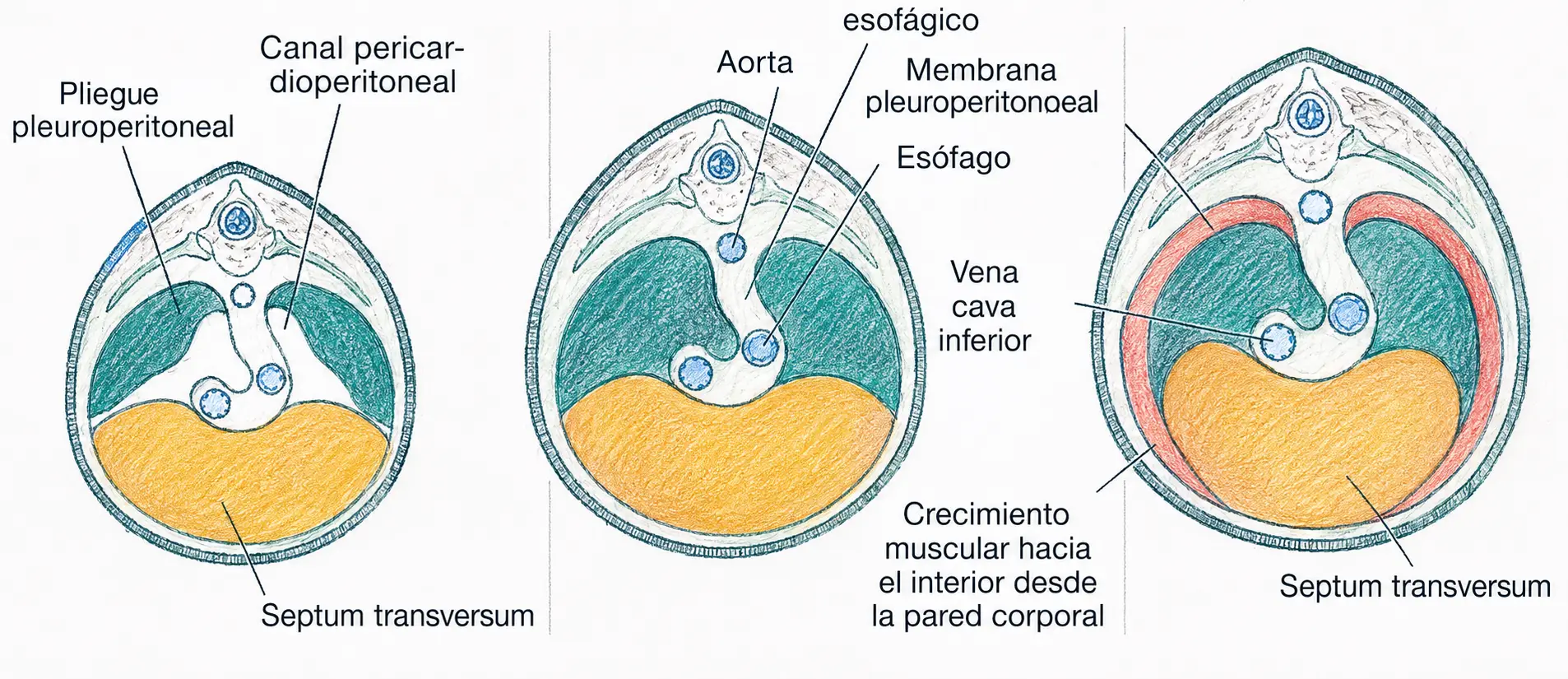
Cuando los pulmones crecen, ocupan progresivamente el espacio entre la somatopleura dorsal y los pliegues pleuropericárdicos, lo que separa definitivamente la futura cavidad pleural de la cavidad pericárdica.
El septum transversum, presente desde estadios iniciales del desarrollo, constituye el tendón central o centro frénico del diafragma. Sobre esta base se completa el cierre de la cavidad celómica en tres compartimentos definitivos: dos cavidades pleurales, la cavidad pericárdica y la cavidad peritoneal.
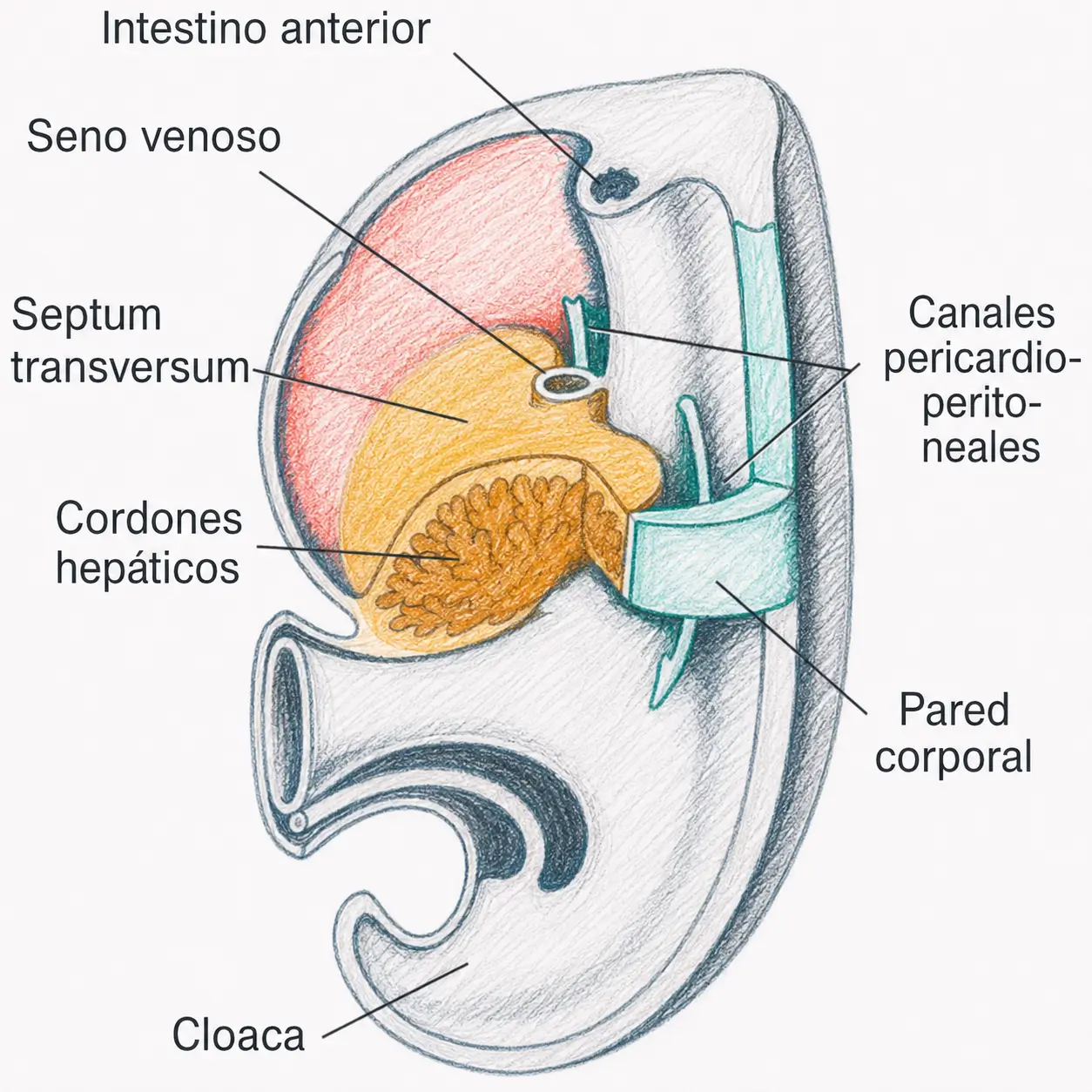
El diafragma completo resulta de la fusión de cuatro componentes distintos, lo que explica por qué es una estructura con tendencia a defectos de fusión:
- El centro frénico (septum transversum).
- El mesoesófago (que aporta el cierre alrededor del esófago).
- Las membranas pleuroperitoneales laterales.
- La musculatura procedente de los somitas cervicales y occipitales.
La masa muscular del propio diafragma no se origina en el propio tórax: procede de somitas cervicales (nivel C4-C5 aproximadamente) que descienden arrastrando su inervación. Por eso el nervio frénico, que inerva el diafragma, nace en la columna cervical y no en la torácica.
Los somitas lumbares forman in situ el declive muscular posterior del diafragma, junto a la aorta, dando lugar a los pilares diafragmáticos.
Precisamente por depender de la fusión correcta de cuatro estructuras independientes, es una de las regiones del desarrollo con más malformaciones clínicamente relevantes.
La más frecuente de todas es la hernia diafragmática congénita de Bochdalek. Se produce cuando la membrana pleuroperitoneal no llega a fusionarse con el centro frénico, dejando un defecto posterolateral en el diafragma, casi siempre en el lado izquierdo. A través de ese defecto, las vísceras abdominales (intestino, y en casos graves también el hígado o el bazo) ascienden a la cavidad torácica durante la vida fetal.
En la hernia de Bochdalek, las vísceras herniadas comprimen el pulmón en desarrollo desde etapas tempranas, antes de que complete sus periodos de maduración. El resultado es hipoplasia pulmonar, con frecuencia bilateral aunque la hernia sea unilateral, que es la verdadera causa de mortalidad en estos recién nacidos, más que la propia hernia.
La hernia de Morgagni es mucho menos frecuente y tiene un mecanismo distinto. Se produce por un hiato anómalamente amplio entre el esternón y el diafragma, en la zona por donde discurre fisiológicamente la arteria mamaria interna (el hiato xifoideo). Suele ser paraesternal derecha, de curso más silente clínicamente que la de Bochdalek, y a menudo se detecta de forma incidental en la vida adulta.
La diferencia entre ambas hernias no es solo la localización (posterolateral en Bochdalek, paraesternal en Morgagni), sino el mecanismo: Bochdalek es un fallo de fusión de una membrana que debería haberse cerrado; Morgagni es un hiato fisiológico que no se estrecha lo suficiente.
Un tercer tipo de defecto, mecánicamente distinto de los dos anteriores porque no implica un orificio nuevo sino la distensión de uno ya existente, es la hernia de hiato. En este caso, el hiato esofágico (por donde el esófago atraviesa fisiológicamente el diafragma para llegar al estómago) se distiende más de lo normal, permitiendo que parte del estómago ascienda hacia el tórax.
Resumen para diferenciar las tres hernias diafragmáticas por su localización:
(1) Bochdalek → posterolateral (fallo de la membrana pleuroperitoneal).
(2) Morgagni → paraesternal, hiato xifoideo (hiato anómalamente amplio).
(3) Hernia de hiato → centro del diafragma, alrededor del esófago (distensión del hiato esofágico fisiológico).
