1.1. Introducción a la gametogénesis
La reproducción sexual exige que los gametos aporten exactamente la mitad de la dotación cromosómica del individuo. Si los gametos fueran diploides, la fecundación duplicaría el número de cromosomas en cada generación. La solución biológica a este problema es la gametogénesis: el proceso mediante el cual células germinales diploides (2n = 46 cromosomas) se transforman en gametos haploides (n = 23 cromosomas) a través de la meiosis.
Gametogénesis: conjunto de procesos de proliferación, diferenciación y maduración celular que transforman células germinales diploides en gametos haploides. En el varón recibe el nombre de espermatogénesis y en la mujer de oogénesis.
En el varón, la espermatogénesis comienza en la pubertad y es un proceso continuo que se prolonga durante toda la vida adulta. En la mujer, la oogénesis comienza antes del nacimiento y sigue una dinámica radicalmente distinta, con largos periodos de arresto meiótico. El T2 de esta asignatura desarrolla la oogénesis en detalle.
La meiosis como mecanismo de división celular, incluyendo sus fases (leptoteno, cigoteno, paquiteno, diploteno, diacinesis, metafase I, etc.) y sus consecuencias genéticas (recombinación, segregación independiente), se estudia en profundidad en Biología Celular (→ Biología, T21-T22: división celular, la meiosis).
Las células germinales tienen un origen extragonadal: se originan en el epiblasto del embrión y migran al esbozo gonadal durante la 5ª-6ª semana del desarrollo. Este origen y la migración de las células germinales primordiales se tratan en el contexto del desarrollo del aparato urogenital (→ T22 de esta asignatura).
1.2. Organización del testículo
Para entender la espermatogénesis es imprescindible conocer el soporte estructural donde ocurre. El testículo contiene dos compartimentos funcionales: los túbulos seminíferos, donde tiene lugar la espermatogénesis, y el tejido intersticial, donde residen las células de Leydig.
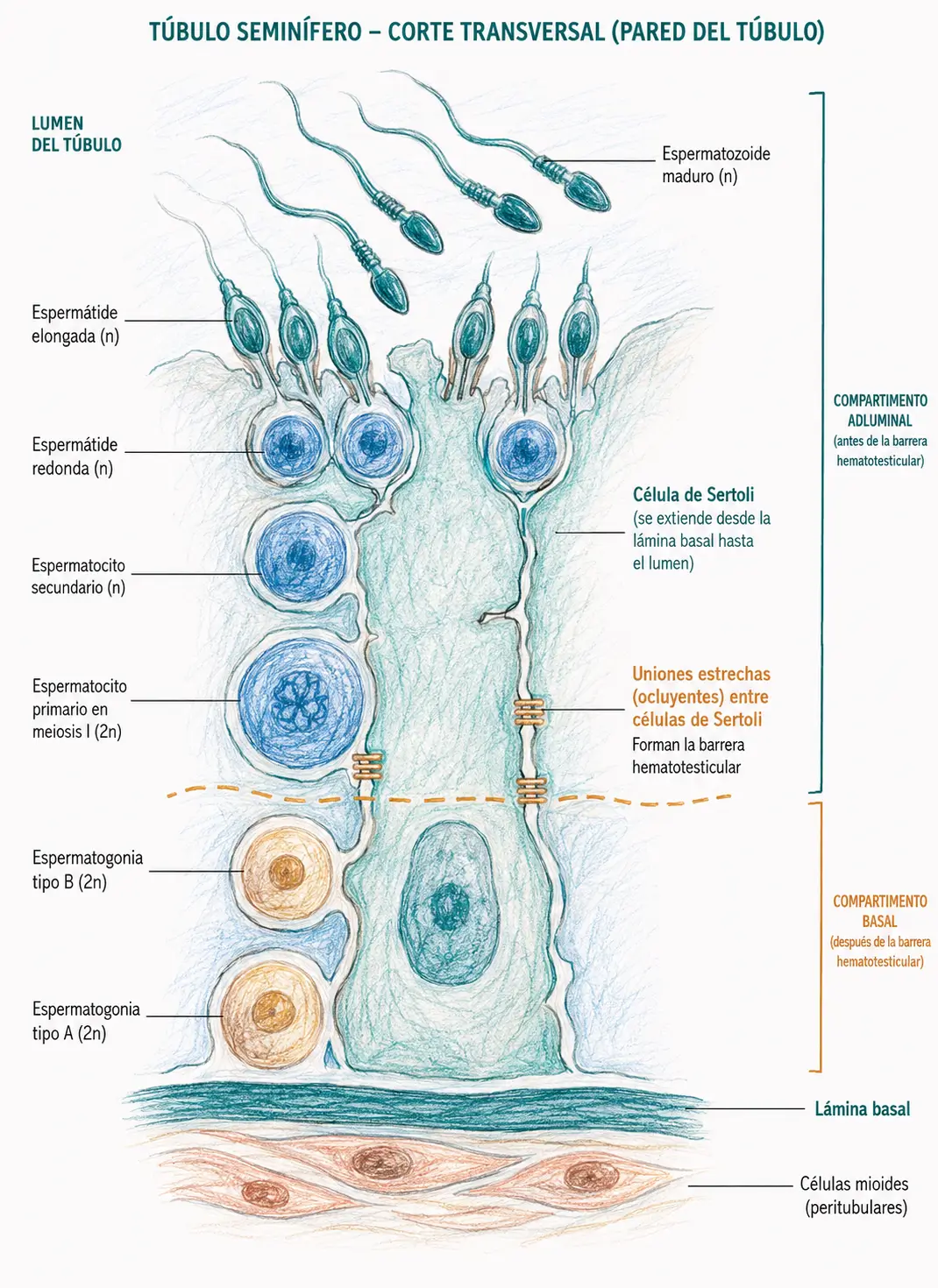
Los túbulos seminíferos son tubos muy replegados (con una longitud total de varios cientos de metros en ambos testículos) cuya pared está formada por dos tipos celulares: las células de Sertoli y las células germinales en distintos estadios de maduración.
Las células de Sertoli son células somáticas no germinales que constituyen el andamiaje del epitelio seminífero. Se extienden desde la membrana basal hasta la luz del túbulo y son imprescindibles para la supervivencia y maduración de las células germinales.
Las células de Sertoli forman entre sí uniones ocluyentes que dividen el epitelio seminífero en dos compartimentos:
- El compartimento basal, donde residen las espermatogonias y los espermatocitos primarios en estadios tempranos.
- El compartimento adluminal, donde se completa la meiosis y ocurre la espermiogénesis.
Este tabique celular constituye la barrera hematotesticular.
La barrera hematotesticular aísla las células germinales en meiosis del sistema inmunitario. Los espermatocitos expresan antígenos propios de la meiosis que serían reconocidos como extraños si entraran en contacto con los linfocitos. La ruptura de esta barrera puede causar infertilidad autoinmune.
Las células de Leydig (o células intersticiales) se sitúan en el tejido conectivo entre los túbulos seminíferos. Su función principal es sintetizar y secretar testosterona bajo el estímulo de la LH hipofisaria. La testosterona es el andrógeno local imprescindible para que la espermatogénesis progrese.
La morfología microscópica del testículo, incluyendo la descripción del epitelio seminífero y el tejido intersticial al microscopio óptico y electrónico, se abordará en Histología Especial (→ Histología Especial: gónada masculina).
1.3. Espermatocitogénesis
La espermatocitogénesis es la primera fase de la espermatogénesis. Comprende desde la proliferación de las espermatogonias hasta la formación de las espermátidas haploides mediante dos divisiones meióticas sucesivas.
1.3.1. Las espermatogonias
Las espermatogonias son las células germinales más inmaduras del epitelio seminífero. Residen en el compartimento basal, en contacto con la membrana basal del túbulo. Se dividen por mitosis de forma continua, asegurando el mantenimiento de la reserva germinal a lo largo de toda la vida reproductiva del varón.
Se distinguen dos tipos funcionales de espermatogonias.
- Las espermatogonias tipo A actúan como células madre: una de las células hijas se autorrenueva, mientras la otra entra en el ciclo de diferenciación.
- Las espermatogonias tipo B son las que se diferenciarán irreversiblemente hacia espermatocitos primarios.
Las espermatogonias tipo A garantizan que la espermatogénesis sea un proceso continuo e inagotable. Si todas las espermatogonias entraran simultáneamente en meiosis, la reserva germinal se agotaría.
1.3.2. Espermatocitos primarios y meiosis I
Las espermatogonias tipo B se diferencian en espermatocitos primarios (2n, con ADN replicado = 4c). Estos cruzan la barrera hematotesticular hacia el compartimento adluminal e inician la meiosis I, la división reductora.
La profase I de la meiosis es la etapa más larga y compleja: dura varios días y comprende los estadios de leptoteno, cigoteno, paquiteno, diploteno y diacinesis. Durante el paquiteno se produce el sobrecruzamiento (crossing-over) entre cromátidas no hermanas de cromosomas homólogos, generando diversidad genética. Tras la división, cada espermatocito primario origina dos espermatocitos secundarios (n, 2c).
1.3.3. Espermatocitos secundarios y meiosis II
Los espermatocitos secundarios son de vida muy corta: entran rápidamente en la meiosis II, la división ecuacional. Esta división es similar a una mitosis: separa las cromátidas hermanas sin reducir de nuevo el número de cromosomas. Cada espermatocito secundario origina dos espermátidas (n, 1c).
El resultado global de la espermatocitogénesis es que cada espermatocito primario produce cuatro espermátidas haploides, todas ellas con igual potencial de convertirse en espermatozoides funcionales.
| Tipo celular | Ploídía / contenido ADN | División | Resultado |
|---|---|---|---|
| Espermatogonia tipo B | 2n / 2c | Mitosis | Espermatocito primario |
| Espermatocito primario | 2n / 4c | Meiosis I (reductora) | 2 espermatocitos secundarios |
| Espermatocito secundario | n / 2c | Meiosis II (ecuacional) | 2 espermátidas |
| Espermátida | n / 1c | Sin división (diferenciación) | Espermatozoide |
1.4. Espermiogénesis
La espermiogénesis es la segunda fase de la espermatogénesis. No implica ninguna división celular: es una transformación morfológica profunda de la espermátida redonda en un espermatozoide estructuralmente maduro y apto para el movimiento.
Los cambios fundamentales que ocurren durante la espermiogénesis son cuatro:
- Formación del acrosoma. El aparato de Golgi sintetiza vesículas que se fusionan y forman la vesícula acrosómica, que se extiende sobre el polo anterior del núcleo. El acrosoma es un orgánulo especializado que contiene enzimas hidrolíticas (principalmente hialuronidasas y acrosinas). Su contenido se libera durante la reacción acrosómica, cuando el espermatozoide contacta con la zona pelúcida del ovocito, y es imprescindible para penetrarla.
- Condensación del núcleo. Las histonas se sustituyen progresivamente por proteínas específicas llamadas protaminas, que compactan el ADN de forma mucho más eficiente. El núcleo se aplana y se desplaza hacia el polo anterior de la célula. Esta compactación extrema protege el material genético paterno durante su tránsito hasta el ovocito.
- Formación del flagelo. Uno de los centriolos se diferencia en el cuerpo basal del flagelo, a partir del cual crece el axonema (estructura 9+2 de microtúbulos). El flagelo proporciona la motilidad progresiva necesaria para alcanzar el ovocito.
- Eliminación del citoplasma residual. La mayor parte del citoplasma de la espermátida se elimina en forma de cuerpo residual, que es fagocitado por las células de Sertoli. Esta pérdida de citoplasma es necesaria para reducir al mínimo el volumen celular y optimizar la hidrodinámica del espermatozoide.
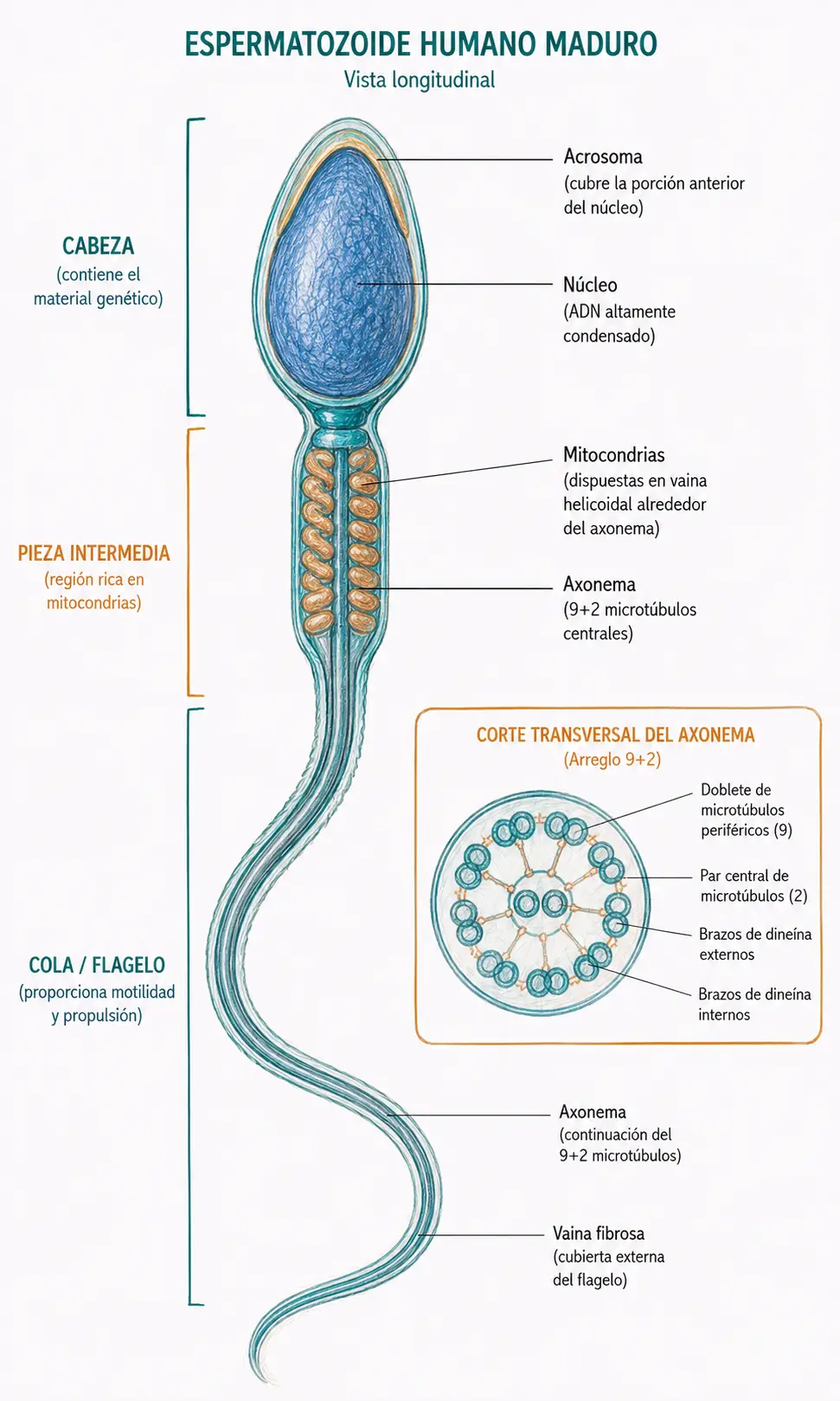
La estructura del espermatozoide maduro se divide en:
- Cabeza (núcleo condensado cubierto por el acrosoma).
- Pieza intermedia (mitocondrias dispuestas en espiral alrededor del axonema, que proporcionan la energía para el movimiento). La pieza intermedia es la fuente de ATP para la motilidad espermática.
- Cola o flagelo (axonema 9+2).
Al final de la espermiogénesis, los espermatozoides son liberados a la luz del túbulo seminífero en un proceso denominado espermiación. En este momento, el espermatozoide es morfológicamente maduro pero todavía inmóvil y sin capacidad fecundante.
Las células germinales que comparten meiosis no se separan completamente: permanecen conectadas por puentes citoplasmáticos hasta el final de la espermiogénesis. Esto sincroniza el desarrollo y permite que moléculas sintetizadas en células con dotación cromosómica XY se compartan con células que solo tienen X o solo Y. Tras la espermiación, los puentes se rompen.
1.5. Maduración en el epidídimo y capacitación
Al ser liberados a la luz tubular, los espermatozoides son trasladados pasivamente hacia el epidídimo, un tubo muy enrollado situado en la cara posterior del testículo. Durante su tránsito por el epidídimo (que dura aproximadamente 12 días en el humano), los espermatozoides adquieren motilidad progresiva y la capacidad de reconocer y unirse a la zona pelúcida. Este proceso de maduración epidídimaria es imprescindible: los espermatozoides directamente extraídos del testículo son incapaces de fecundar.
La capacitación es un proceso distinto, posterior, que ocurre en el tracto genital femenino (cuello uterino, útero, trompas de Falopio). Consiste en cambios bioquímicos de la membrana plasmática del espermatozoide, incluyendo:
- La eliminación del colesterol de membrana.
- La modificación de proteínas de superficie, que hiperactivarán su motilidad y prepararán la reacción acrosómica.
La capacitación requiere entre 5 y 7 horas.
La capacitación no es lo mismo que la maduración epidídimaria. La maduración ocurre en el epidídimo y otorga motilidad progresiva. La capacitación ocurre en el tracto genital femenino y es el paso funcional final que hace al espermatozoide competente para la fecundación.
1.6. Regulación hormonal de la espermatogénesis
La espermatogénesis está bajo control del eje hipotálamo-hipófisis-testículo. El hipotálamo secreta GnRH (hormona liberadora de gonadotropinas) de forma pulsátil, que actúa sobre la adenohipófisis estimulando la liberación de dos gonadotropinas: FSH (hormona foliculoestimulante) y LH (hormona luteinizante).
- La FSH actúa directamente sobre los receptores de las células de Sertoli, estimulando la producción de ABP (proteína ligadora de andrógenos) y de inhibina, y potenciando el soporte nutricional a las células germinales.
- La LH actúa sobre las células de Leydig, estimulando la síntesis y secreción de testosterona. La testosterona es el regulador local más importante de la espermatogénesis: concentraciones altas de testosterona dentro del túbulo (mantenidas por la ABP de Sertoli) son imprescindibles para que la meiosis y la espermiogénesis progresen con normalidad.
El sistema dispone de retroalimentación negativa: la testosterona inhibe la secreción de LH (y de GnRH), y la inhibina producida por las células de Sertoli inhibe selectivamente la secreción de FSH.
El conocimiento de este eje hormonal tiene aplicación directa en clínica. El hipogonadismo hipogonadotropo (déficit de GnRH, FSH o LH) produce azoospermia tratable con gonadotropinas exógenas. Por el contrario, la administración exógena de testosterona o anabolizantes androgénicos suprime la GnRH y las gonadotropinas endógenas por retroalimentación negativa, inhibiendo la espermatogénesis y causando infertilidad. Este es el mecanismo por el que el dopaje con esteroides anabolizantes produce atrofia testicular e infertilidad.
1.7. Duración y rendimiento de la espermatogénesis
El ciclo completo de la espermatogénesis, desde espermatogonia tipo B hasta la espermiación, dura aproximadamente 74 días en el humano. A esto hay que sumar el tránsito por el epidídimo (12 días), lo que significa que el proceso completo hasta tener espermatozoides maduros y capacitados tarda unos 3 meses.
El rendimiento es extraordinario: en condiciones normales, el testículo produce varios millones de espermatozoides por día. Se estima que un eyaculado humano contiene entre 40 y 600 millones de espermatozoides.
La duración del ciclo espermatogénico tiene implicaciones clínicas relevantes. Los agentes gonadotóxicos (quimioterapia, radioterapia, fiebre alta prolongada) que dañan el epitelio seminífero provocan una caída en el recuento espermático que no se manifiesta hasta 2-3 meses después de la exposición. Del mismo modo, la recuperación tras la supresión de un tratamiento gonadotóxico también tarda varios meses.
