3.1. Introducción: la citogenética y su relevancia clínica
A mediados del siglo XX, la confluencia entre la biología celular, la genética y la citología dio lugar a una nueva disciplina: la citogenética. Su objeto de estudio son los cromosomas, su estructura, su número y las alteraciones que pueden sufrir, así como la relación entre esas alteraciones y la clínica de los pacientes.
La relevancia médica de la citogenética es inmediata: las anomalías cromosómicas explican una parte muy significativa de los abortos espontáneos del primer trimestre, algunas malformaciones fetales y síndromes reconocibles desde el nacimiento. Saber interpretar un cariotipo es, por tanto, una competencia clínica básica.
Levan y Tjio establecieron en 1956 el número de cromosomas de la especie humana: 46. El impulso definitivo llegó en 1959, cuando Lejeune demostró que el síndrome de Down se asocia a una trisomía del cromosoma 21. Estas dos fechas son referencia habitual en preguntas de examen.
Hasta 1956 el número de cromosomas humanos era desconocido con certeza: recuentos previos arrojaban cifras tan dispares como 29, 48 o 80. El problema era técnico: en la placa metafásica los cromosomas aparecen superpuestos y son difíciles de contar individualmente. Levan y Tjio lo resolvieron combinando dos avances metodológicos: el cultivo de tejidos in vitro y la técnica del choque hipotónico, que hincha las células y separa físicamente los cromosomas. El mismo año, Ford y Hamerton confirmaron el número de 46 analizando espermatogonias en biopsias testiculares, y establecieron además la existencia de dos gonosomas distintos, X e Y.
El verdadero arranque de la citogenética clínica ocurrió tres años después. Lejeune demostró que los niños con síndrome de Down presentaban 47 cromosomas: un cromosoma 21 supernumerario. Era la primera vez que una anomalía numérica cromosómica se relacionaba directamente con una enfermedad humana, lo que abrió la puerta al estudio sistemático de las anomalías cromosómicas como causa de patología.
3.2. Obtención del cariotipo: metodología paso a paso
El cariotipo es la imagen que se obtiene cuando los cromosomas de una célula en metafase se ordenan por parejas de homólogos, de mayor a menor tamaño y siguiendo criterios internacionales establecidos. Para obtenerlo se necesitan células en división activa que puedan detenerse en metafase para analizar sus cromosomas.
La muestra de elección es la sangre periférica. Los linfocitos maduros no se dividen espontáneamente una vez diferenciados, pero en 1960 Moorhead descubrió que podían reactivarse in vitro. Este hallazgo fue decisivo: convertir el análisis citogenético en una técnica de rutina a partir de una muestra de fácil obtención.
Protocolo del cariotipo a partir de sangre periférica:
- Extracción de sangre periférica en condiciones de esterilidad, con heparina para prevenir la coagulación.
- Cultivo en medio RPMI suplementado con suero bovino fetal y antibióticos, durante 72 horas.
- Adición de fitohemaglutinina: lectina que se une a la membrana de los linfocitos y los induce a desdiferenciarse en linfoblastos capaces de dividirse. En las primeras 24 horas los linfocitos se transforman en linfoblastos, que proliferan estimulados por interleuquina 2.
- A las 70–71 horas: adición de colchicina, que bloquea la polimerización de microtúbulos y destruye el huso mitótico. Las células quedan detenidas en metafase, con los cromosomas condensados e individualizados.
- Choque hipotónico con solución de KCl: las células se hinchan por ósmosis y los cromosomas se dispersan en el interior celular.
- Fijación con fijador de Carnoy (metanol/ácido acético): preserva la morfología cromosómica.
- Extensiones sobre portaobjetos, tinción y análisis.
La colchicina detiene la mitosis en metafase, no en anafase. Actúa impidiendo la polimerización de la tubulina, de modo que el huso acromático no se forma y las cromátidas no pueden migrar. El resultado es la acumulación de células con cromosomas perfectamente condensados e individualizados, ideales para el análisis.
Además de sangre periférica, se pueden utilizar otras fuentes celulares según el contexto clínico: células del líquido amniótico o de vellosidades coriales en el diagnóstico prenatal, células de médula ósea en hematología, o células de biopsia testicular cuando interesa estudiar específicamente la meiosis.
Una vez obtenidas las metafases, los cromosomas se cuentan, se fotografían y se ordenan. Actualmente este proceso está automatizado mediante programas informáticos de análisis de imagen.
3.3. Clasificación de los cromosomas humanos: grupos A–G
El cariotipo humano en células somáticas contiene 46 cromosomas: 22 pares de autosomas y 1 par de gonosomas (XX en la mujer, XY en el hombre). Para ordenarlos se utilizan tres criterios:
- Tamaño.
- Posición del centrómero.
- Presencia de constricciones secundarias o satélites.
Con estos parámetros se establecen siete grupos, designados con las letras A a G.
Posición del centrómero: determina la morfología del cromosoma. Si el centrómero divide el cromosoma en dos brazos iguales se llama metacéntrico; si los divide de forma desigual pero no extrema, submetacéntrico; si está muy desplazado hacia uno de los extremos, acrocéntrico.
| Grupo | Cromosomas | Características |
|---|---|---|
| A | 1, 2, 3 | Los más grandes del cariotipo. Cr. 1 y 3 metacéntricos; cr. 2 submetacéntrico. |
| B | 4, 5 | Grandes, muy submetacéntricos. Difíciles de diferenciar entre sí. |
| C | 6–12, X | Grupo más numeroso y complejo. Submetacéntricos de tamaño mediano. El cromosoma X es submetacéntrico, de tamaño intermedio entre el 6 y el 7. |
| D | 13, 14, 15 | Acrocéntricos de tamaño mediano. Presentan constricciones secundarias con pequeños satélites donde se localizan los genes del ARN ribosómico (organizadores nucleolares, NOR). |
| E | 16, 17, 18 | Tamaño pequeño-mediano. Cr. 16 casi metacéntrico; cr. 17 submetacéntrico; cr. 18 muy submetacéntrico. |
| F | 19, 20 | Pequeños, metacéntricos. Se denominan "pequeñas cruces". Casi indistinguibles entre sí. |
| G | 21, 22, Y | Los más pequeños. Cr. 21 y 22 acrocéntricos con satélites NOR. Cr. Y submetacéntrico, de tamaño variable, con cromátidas muy paralelas y zona distal de heterocromatina constitutiva. |
El cromosoma 21 es en realidad el más pequeño del cariotipo humano; el 22 es morfológicamente algo mayor. Sin embargo, el sistema de numeración actual lo mantiene en esa posición por razones históricas: fue el primero en asociarse a un síndrome clínico concreto (Down) y protagonizó el arranque de la citogenética clínica moderna. Cambiarlo ahora supondría alterar toda la nomenclatura acumulada durante décadas.
3.4. Técnicas de bandeado
Con el cariotipo convencional los cromosomas del mismo grupo son difíciles de distinguir entre sí. A partir de 1970 se desarrollaron las técnicas de bandeado: tratamientos físicos o químicos que generan patrones alternantes de bandas oscuras y claras característicos e individuales para cada cromosoma. Estas técnicas permiten identificar cada cromosoma con certeza, caracterizarlo por regiones y relacionar alteraciones concretas con enfermedades.
Existen dos categorías:
- Técnicas que bandean todo el cromosoma (Q, G, R).
- Técnicas que marcan regiones específicas (C, NOR, T).
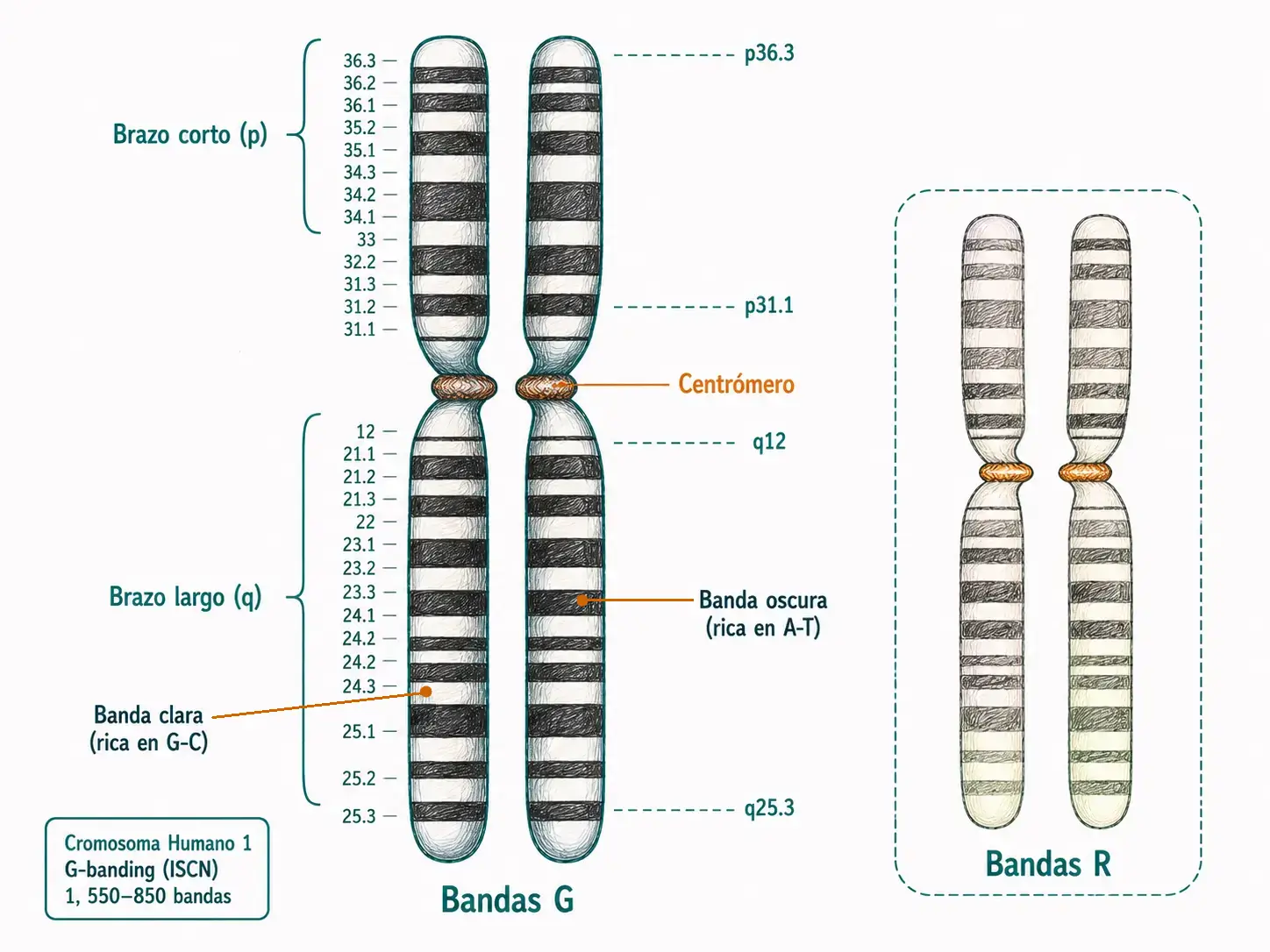
3.4.1. Bandas de todo el cromosoma
Las bandas Q (quinacrina) fueron las primeras en desarrollarse. Se obtienen tiñendo con quinacrina y observando con microscopio de fluorescencia ultravioleta. El resultado es un patrón de bandas más o menos fluorescentes específico de cada cromosoma. Su ventaja es la rapidez; su inconveniente, que la fluorescencia se pierde en menos de un minuto.
Las bandas G (Giemsa) son el estándar actual para el cariotipo de rutina. Se obtienen tratando los cromosomas con tripsina (una solución proteolítica) y tiñendo posteriormente con Giemsa. Las regiones oscuras corresponden a zonas de replicación tardía, ricas en adenina y timina. El patrón resultante permite individualizar todos los cromosomas y reconocerlos inequívocamente.
Las bandas R son el negativo de las bandas G: lo que aparece oscuro en G aparece claro en R, y viceversa. Se obtienen tratando los cromosomas con solución salina caliente antes de Giemsa. Tiñen preferentemente las regiones de replicación temprana, ricas en guanina y citosina. Son muy utilizadas en Francia y resultan útiles cuando las regiones teloméricas necesitan mejor definición.
Bandas G y bandas R son complementarias, no equivalentes. Las bandas G marcan heterocromatina tardía (AT-rich, oscura); las bandas R marcan eucromatina temprana (GC-rich, oscura). Confundirlas lleva a interpretar erróneamente la localización de una deleción o una translocación.
El cariotipo en metafase ofrece alrededor de 400 bandas en haploide. Si se trabaja con cromosomas prometafásicos, más elongados, se pueden resolver hasta 2.000 bandas, lo que aumenta la resolución para detectar alteraciones pequeñas.
3.4.2. Técnicas que marcan regiones específicas
Las bandas C tiñen selectivamente la heterocromatina constitutiva: los centrómeros, las regiones heterocromatínicas de los cromosomas 1, 9 y 16, los brazos cortos de los acrocéntricos y el brazo largo distal del cromosoma Y.
Las bandas NOR (nucleolar organizer regions) tiñen los satélites de los cromosomas acrocéntricos (13, 14, 15, 21 y 22), donde se localizan los genes del ARN ribosómico 18S y 28S.
Las bandas T marcan los telómeros y permiten detectar pérdidas teloméricas.
3.4.2. Hibridación in situ fluorescente (FISH)
La hibridación in situ fluorescente (FISH, del inglés Fluorescence In Situ Hybridization) supera las limitaciones de resolución del bandeado convencional. Utiliza sondas de ADN marcadas con fluorocromos que hibridan específicamente con regiones o genes concretos del cromosoma. Permite detectar microdeleciones, amplificaciones génicas y reordenamientos que no serían visibles con el bandeado clásico.
La FISH tiene aplicación clínica directa en el diagnóstico de síndromes de microdeleción (síndrome de DiGeorge por deleción 22q11, síndrome de Williams por deleción 7q11), en el seguimiento de trasplantes de médula ósea y en la detección de translocaciones oncogénicas como la BCR-ABL de la leucemia mieloide crónica.
3.5. Nomenclatura del cariotipo
La nomenclatura citogenética está estandarizada a nivel internacional mediante el sistema ISCN (International System for Human Cytogenomic Nomenclature). La fórmula básica se construye indicando primero el número total de cromosomas, seguido de una coma y la constitución de los gonosomas.
Un cariotipo femenino normal se escribe 46,XX y un cariotipo masculino normal 46,XY.
Cualquier desviación de esta fórmula indica una anomalía: la adición de un cromosoma se señala con el signo más (+) y la pérdida con el signo menos (–), seguidos del número del cromosoma afectado. Así, 47,XY,+21 describe el cariotipo de un varón con trisomía 21 (síndrome de Down) y 45,X describe el cariotipo de la monosomía del cromosoma X (síndrome de Turner).
Las anomalías numéricas y estructurales del cariotipo, así como su mecanismo de producción durante la meiosis, se estudian en el Tema 4 · Anomalías numéricas de los cromosomas y el Tema 5 · Anomalías estructurales de los cromosomas
