10.1. Implantación completa y reacción decidual
Al inicio de la segunda semana, el blastocisto ha iniciado la implantación en el endometrio pero todavía no está completamente enterrado. Durante los días 8 a 12, el sincitiotrofoblasto continúa su expansión invasiva y el blastocisto se introduce progresivamente en el espesor del endometrio hasta quedar completamente recubierto por el epitelio endometrial, que se cierra por encima de él. En este momento se completa la implantación intersticial.
La invasión del sincitiotrofoblasto provoca una respuesta local en el estroma endometrial denominada reacción decidual: las células estromales se transforman en células deciduales, redondeadas y grandes, ricas en glucógeno y lípidos. Esta transformación afecta inicialmente al estroma circundante al blastocisto y se extiende progresivamente a todo el endometrio bajo la influencia de la progesterona del cuerpo lúteo.
La decidua es el endometrio modificado por la reacción decidual durante el embarazo. Se divide en tres regiones según su relación con el embrión: decidua basal (entre el embrión y el miometrio, que formará parte de la placenta), decidua capsular (que cubre el embrión por el lado de la cavidad uterina) y decidua parietal (resto del endometrio uterino).
La reacción decidual cumple varias funciones: proporciona nutrientes al embrión en implantación, limita la profundidad de la invasión trofoblástica y modula la respuesta inmune materna para tolerar el semialógrafo que representa el embrión.
El embrión es genéticamente distinto de la madre (contiene genes paternos) y debería ser reconocido como extraño por el sistema inmune materno. La tolerancia inmunológica del embarazo es un fenómeno activo y complejo que implica la inducción de células T reguladoras, la expresión de moléculas HLA no clásicas por el trofoblasto y la acción inmunomoduladora de la progesterona y la hCG. Su fallo puede contribuir a la pérdida gestacional recurrente.
10.2. Diferenciación del trofoblasto
Durante la segunda semana, el trofoblasto se diferencia en dos capas con funciones distintas que se mantienen durante todo el desarrollo placentario.
El citotrofoblasto es la capa interna, formada por células individuales con capacidad mitótica. Actúa como reservorio proliferativo: sus células se dividen y migran hacia el sincitiotrofoblasto, fusionándose con él y aportando nuevos núcleos. El citotrofoblasto no tiene capacidad invasiva directa por sí mismo.
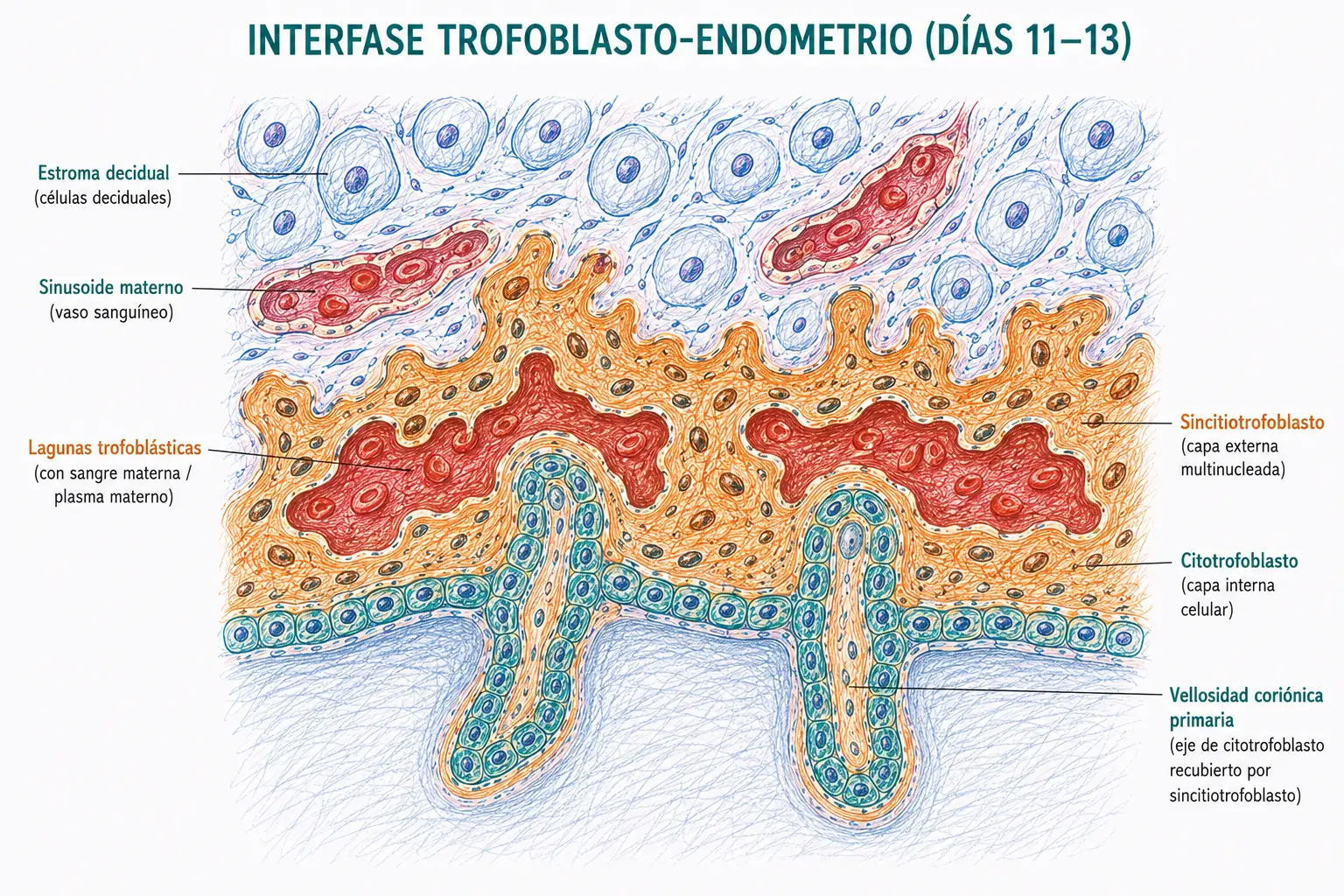
El sincitiotrofoblasto es la capa externa, formada por la fusión de células del citotrofoblasto que origina un sincitio multinucleado sin límites celulares definidos. Tiene potente capacidad invasiva: secreta metaloproteasas de matriz y otras enzimas proteolíticas que digieren el colágeno y la fibronectina del estroma endometrial. Además, el sincitiotrofoblasto es la principal fuente de hCG durante las primeras semanas del embarazo.
La hCG producida por el sincitiotrofoblasto actúa sobre el cuerpo lúteo del ovario como sustituto de la LH, manteniéndolo activo y estimulando la producción continua de progesterona. Sin esta señal, el cuerpo lúteo degeneraría hacia el día 14, la progesterona caería y se produciría la menstruación con pérdida del embrión. La hCG es literalmente la señal de rescate del cuerpo lúteo.
10.3. Lagunas trofoblásticas y circulación uteroplacentaria primitiva
A partir del día 9, el sincitiotrofoblasto erosiona las paredes de los sinusoides maternos del endometrio. La sangre materna comienza a fluir hacia espacios que se forman dentro del propio sincitiotrofoblasto: las lagunas trofoblásticas. Estas lagunas, al principio pequeñas e independientes, se comunican progresivamente entre sí formando una red lacunar.
Al mismo tiempo, desde el lado fetal, el citotrofoblasto prolifera y envía proyecciones hacia el interior del sincitiotrofoblasto: son las vellosidades coriónicas primarias (estadio del día 13-14), formadas inicialmente solo por citotrofoblasto recubierto de sincitiotrofoblasto.
La combinación de las lagunas trofoblásticas llenas de sangre materna y las vellosidades coriónicas que se proyectan hacia ellas establece la circulación uteroplacentaria primitiva: sangre materna bañando las vellosidades trofoblásticas. Este es el precursor de la placenta definitiva, en la que los intercambios materno-fetales tendrán lugar a través de las membranas de las vellosidades.
En este momento todavía no hay vasos fetales en las vellosidades: el intercambio es por difusión simple a través de las capas del trofoblasto. Los vasos sanguíneos fetales no aparecen en las vellosidades hasta la tercera semana, cuando las vellosidades se vascularizarán y se convertirán en vellosidades terciarias.
10.4. Formación del disco germinativo bilaminar
Mientras el trofoblasto lleva a cabo la implantación, la masa celular interna (embrioblasto) se reorganiza para formar el disco germinativo bilaminar. Este proceso comienza hacia el día 8 y se completa hacia el día 14.
Las células del embrioblasto se diferencian en dos capas epiteliales superpuestas que forman un disco plano:
El epiblasto es la capa superior, formada por células cilíndricas altas estrechamente unidas. Está orientado hacia la cavidad amniótica que se forma por encima de él. El epiblasto es la capa más importante del disco bilaminar: de él derivarán, mediante la gastrulación de la tercera semana, las tres hojas germinativas (ectodermo, mesodermo y endodermo) y, por tanto, todos los tejidos del embrión propiamente dicho.
El hipoblasto es la capa inferior, formada por células cúbicas pequeñas. Está orientado hacia la cavidad del blastocele. El hipoblasto no contribuye a los tejidos embrionarios: sus células migran para tapizar la cara interna del citotrofoblasto y formar el saco vitelino primario (o saco vitelino primitivo, también denominado cavidad exocelómica o membrana de Heuser).
Es un error frecuente atribuir al hipoblasto un papel en la formación del embrión. El hipoblasto da lugar exclusivamente a estructuras extraembrionarias: el revestimiento del saco vitelino y parte del mesodermo extraembrionario visceral. Todos los tejidos del embrión, sin excepción, derivan del epiblasto.
10.5. Cavidades extraembrionarias
Durante la segunda semana aparecen tres espacios cavitarios que, aunque no forman parte del embrión propiamente dicho, son estructuras de sostén y nutrición esenciales para su desarrollo.
10.5.1. Cavidad amniótica
Por encima del epiblasto, algunas células del citotrofoblasto se diferencian en amnioblastos y delimitan una cavidad llena de líquido: la cavidad amniótica. Esta cavidad está tapizada por el amnios, una membrana avascular muy delgada. El epiblasto forma el suelo de la cavidad amniótica.
La cavidad amniótica irá creciendo hasta rodear por completo al embrión. El líquido amniótico que la llena protege al embrión de traumatismos mecánicos, mantiene una temperatura constante y permite los movimientos fetales necesarios para el desarrollo musculoesquelético.
10.5.2. Saco vitelino
Por debajo del hipoblasto, las células hipoblásticas proliferan y migran para tapizar la cara interna del citotrofoblasto, formando el saco vitelino primario. Poco después, una segunda oleada de células hipoblásticas desplaza parcialmente al saco vitelino primario, reduciendo su tamaño: el remanente se denomina saco vitelino secundario (o saco vitelino definitivo).
En humanos el saco vitelino no contiene vitelo (nutrientes) como en otros vertebrados: su función principal es ser el primer sitio de hematopoyesis extraembrionaria (producción de células sanguíneas) y albergar las células germinales primordiales que migrarán posteriormente a las gónadas.
El saco vitelino es el primer sitio de hematopoyesis en el embrión humano, a partir de la tercera semana. Las células germinales primordiales (precursoras de los gametos) también residen inicialmente en la pared del saco vitelino antes de migrar a las gónadas. Ambos datos son pregunta frecuente de examen.
10.5.3. Mesodermo extraembrionario y cavidad coriónica
Hacia el día 12-13, entre el trofoblasto y las membranas del amnios y del saco vitelino, aparece una nueva población celular: el mesodermo extraembrionario (también llamado mesodermo del saco vitelino o mesodermo extraembrionario de origen hipoblástico, aunque su origen exacto es debatido). Este mesodermo se dispone como una capa laxa de células que rellena el espacio entre el trofoblasto y las estructuras embrionarias.
Progresivamente, dentro del mesodermo extraembrionario aparecen pequeñas cavidades que confluyen hasta formar un espacio único: la cavidad coriónica o celoma extraembrionario. Esta cavidad separa el mesodermo extraembrionario en dos hojas: la hoja somatopleural (adosada al trofoblasto, que junto a este forma el corion) y la hoja esplacnopleural (adosada al amnios y al saco vitelino).
La cavidad coriónica crece hasta rodear completamente al embrión y sus membranas, excepto en un punto: el pedículo de fijación, una banda de mesodermo extraembrionario que conecta el polo caudal del disco embrionario con el trofoblasto. El pedículo de fijación es el precursor del cordón umbilical.
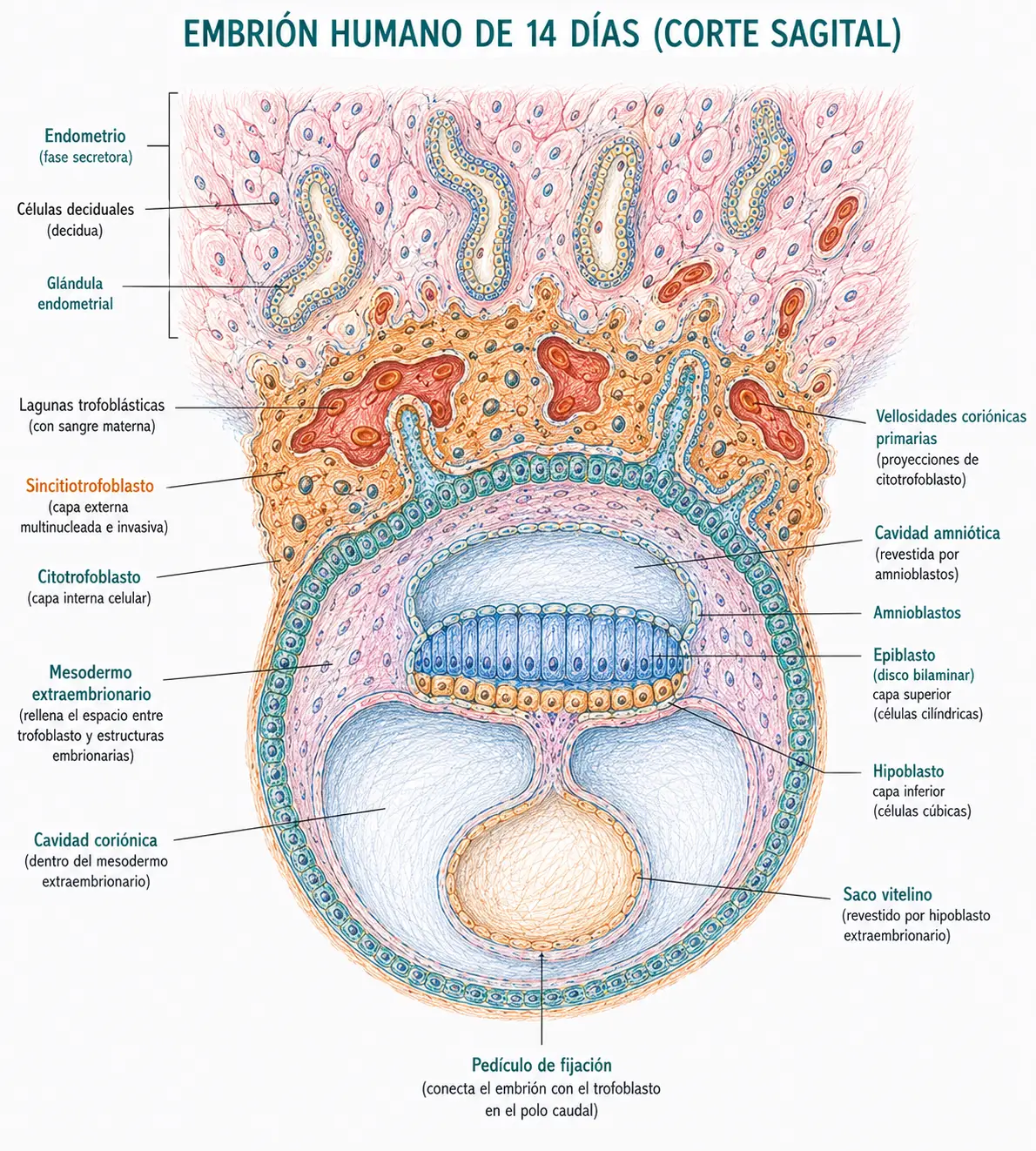
Al final de la segunda semana, la organización espacial del embrión ya tiene su arquitectura básica establecida: el disco bilaminar en el centro, suspendido en la cavidad coriónica, conectado a la pared del trofoblasto únicamente por el pedículo de fijación, con la cavidad amniótica por encima y el saco vitelino por debajo.
10.6. La regla de los dos: el día 14
La segunda semana culmina alrededor del día 14 con una serie de eventos que se describen clásicamente como la regla de los dos: aparecen dos capas del trofoblasto (citotrofoblasto y sincitiotrofoblasto), dos capas del disco embrionario (epiblasto e hipoblasto), dos cavidades (amniótica y saco vitelino), dos hojas del mesodermo extraembrionario (somatopleural y esplacnopleural), y comienza a formarse la línea primitiva que marcará el inicio de la gastrulación.
La regla de los dos es un recurso mnemotécnico clásico para recordar los eventos de la segunda semana: 2 capas trofoblásticas, 2 capas del disco, 2 cavidades (amnios y saco vitelino), 2 hojas del mesodermo extraembrionario. Todo ocurre durante la segunda semana y todo viene de a dos.
Al final del día 14 aparece en la superficie del epiblasto la línea primitiva, una condensación celular en la línea media caudal del disco que marca el inicio de la gastrulación. Con la aparición de la línea primitiva termina el periodo preembrionario y comienza el periodo embrionario propiamente dicho, que se estudia en el Tema 6 · Tercera semana y gastrulación.
La aparición de la línea primitiva en el día 14 tiene relevancia ética y legal en muchas legislaciones: es el límite temporal a partir del cual los embriones humanos no pueden ser mantenidos en cultivo con fines de investigación (la denominada regla de los 14 días). Antes de la línea primitiva no existe un eje corporal definido ni individualidad establecida; es también antes de este momento cuando puede producirse la gemelaridad monocigótica. Varios países están debatiendo ampliar este límite a 28 días a la luz de los avances en cultivo embrionario in vitro.
