11.1. Definición de gastrulación
La gastrulación es uno de los eventos más importantes del desarrollo embrionario. Al comenzar la tercera semana, el embrión es un disco bilaminar de dos capas (epiblasto e hipoblasto) sin ninguna diferenciación regional aparente más allá del eje embrionario-abembrionario. Al terminar la tercera semana, el embrión tiene tres hojas germinativas, un eje cráneo-caudal definido, un eje dorso-ventral establecido y una estructura axial inductora (la notocorda) en su lugar.
Gastrulación: proceso morfogenético que transforma el disco bilaminar en un disco trilaminar mediante la formación de las tres hojas germinativas primarias: ectodermo, mesodermo intraembrionario y endodermo definitivo. Es el inicio del periodo embrionario propiamente dicho y el momento en que se establecen los ejes corporales básicos.
Las tres hojas germinativas son los precursores de todos los tejidos y órganos del organismo adulto:
- Del ectodermo derivarán el sistema nervioso, la epidermis y los órganos de los sentidos.
- Del mesodermo derivarán el aparato locomotor, el aparato cardiovascular, el aparato urogenital y el tejido conjuntivo.
- Del endodermo derivarán el epitelio del tubo digestivo, el aparato respiratorio y las glándulas asociadas al tubo digestivo.
El destino de cada hoja germinativa, es decir, qué tejidos y órganos derivan de cada una, se estudia en detalle en los temas de organogénesis de esta asignatura (del tema 14 al tema 24). La histología de los tejidos derivados de cada hoja se estudia en Histología General y Especial (→ Histología General 34443).
11.2. La línea primitiva
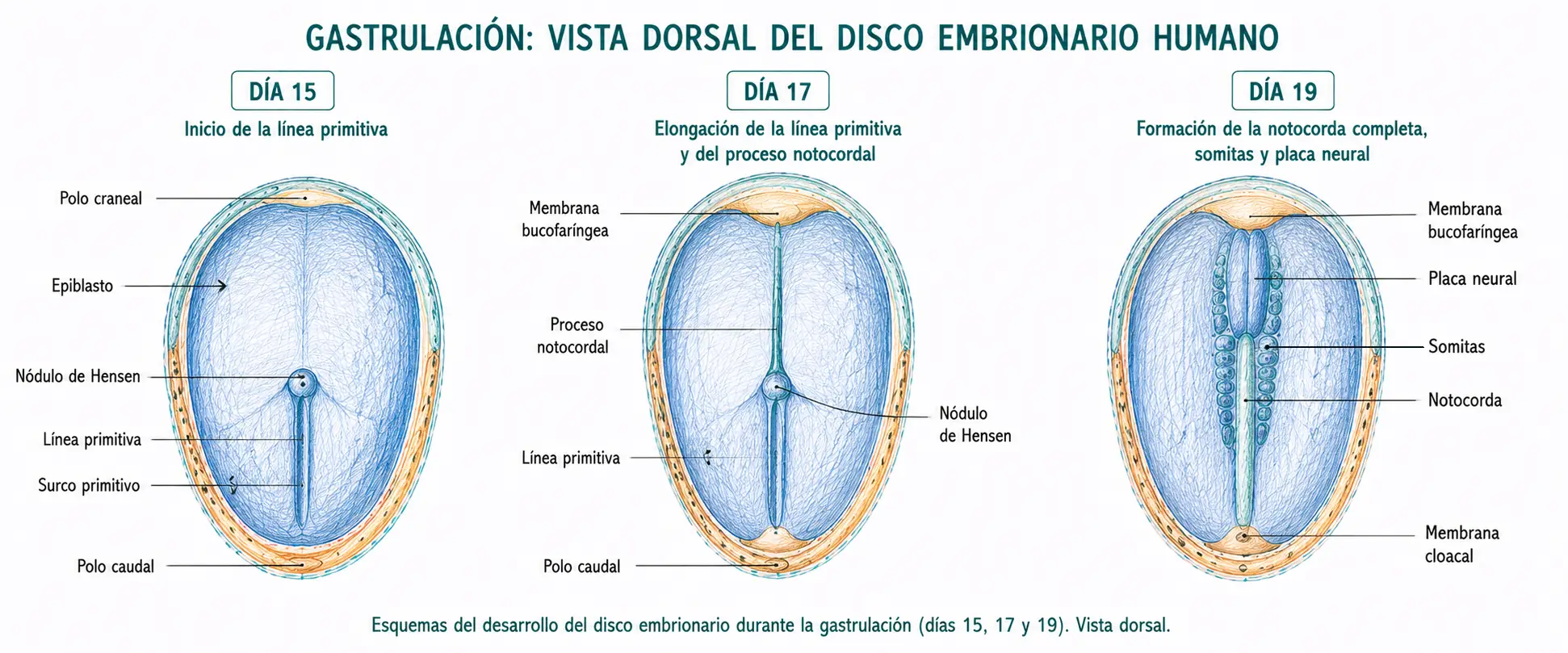
La gastrulación se inicia con la aparición de la línea primitiva en la superficie del epiblasto, hacia el día 14-15. Es una condensación lineal de células epiblásticas en la región caudal y medial del disco embrionario.
La línea primitiva tiene una orientación cráneo-caudal y su aparición es el primer indicador morfológico del eje del embrión: el extremo cefálico de la línea primitiva señala el polo craneal del embrión y su extremo caudal señala el polo caudal. Con su aparición queda también definido el eje izquierda-derecha, ya que la línea es estrictamente medial.
La línea primitiva es el organizador morfogenético central de la gastrulación. Establece simultáneamente tres ejes: el eje cráneo-caudal (su longitud define el eje anterior-posterior), el eje dorso-ventral (las células que se invaginan por ella se dirigen hacia el interior) y el eje izquierda-derecha (su posición estrictamente medial divide el embrión en dos mitades simétricas). Sin línea primitiva no hay gastrulación posible.
En el extremo craneal de la línea primitiva se forma una condensación celular más prominente denominada nódulo de Hensen (o nódulo primitivo). El nódulo de Hensen es funcionalmente distinto del resto de la línea: las células que migran a través de él no formarán mesodermo lateral sino la notocorda, la estructura axial más importante del embrión temprano.
A lo largo de la línea primitiva y del nódulo de Hensen se forma una depresión o surco: el surco primitivo y la foseta primitiva respectivamente. Estas depresiones son los puntos de invaginación de las células epiblásticas durante la gastrulación.
11.3. Movimientos celulares de la gastrulación
La gastrulación implica un proceso coordinado de migración celular denominado ingresión. Las células del epiblasto sufren una transición epitelio-mesenquimal:
- Pierden sus uniones con las células vecinas.
- Adquieren capacidad migratoria.
- Se desplazan hacia la línea primitiva.
Al llegar a la línea primitiva, las células se invaginan (pasan hacia el interior del disco) a través del surco primitivo. Una vez en el interior, migran en tres direcciones:
- Las primeras células en migrar lo hacen profundamente, desplazando y sustituyendo al hipoblasto. Estas células forman el endodermo definitivo, que tapizará el tubo digestivo y el aparato respiratorio. El hipoblasto desplazado queda restringido al revestimiento del saco vitelino.
- Las células que migran posteriormente se interponen entre el ectodermo (epiblasto restante) y el endodermo recién formado, extendiéndose lateralmente en todas las direcciones. Estas células constituyen el mesodermo intraembrionario.
- Las células del epiblasto que no migran a través de la línea primitiva permanecen en la superficie del disco y constituyen el ectodermo.
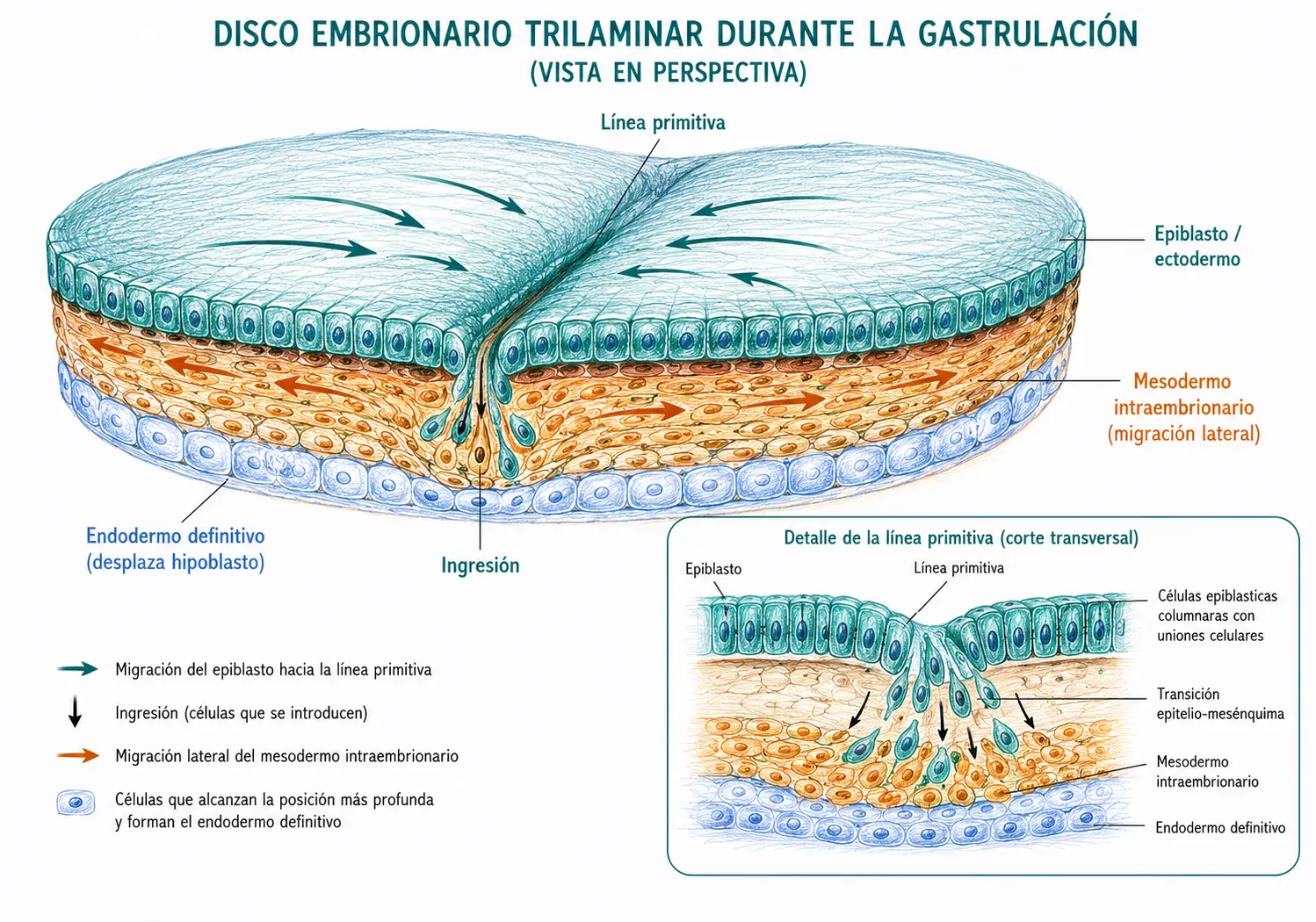
Las tres hojas germinativas derivan exclusivamente del epiblasto. El hipoblasto no contribuye a ningún tejido embrionario: es completamente desplazado por el endodermo de nueva formación durante la gastrulación. Este es uno de los conceptos más contraintuitivos de la embriología temprana y uno de los errores más frecuentes en examen.
11.4. Zonas sin mesodermo intraembrionario
El mesodermo intraembrionario se extiende por prácticamente todo el disco embrionario, pero hay dos regiones donde ectodermo y endodermo están en contacto directo sin mesodermo interpuesto.
- La membrana bucofaríngea es una zona circular en el extremo craneal del disco donde ectodermo y endodermo están fusionados. Marca el futuro extremo anterior del tubo digestivo. Al romperse hacia el final de la cuarta semana, formará la cavidad bucal primitiva (estomodeo) y comunicará el exterior con el intestino anterior.
- La membrana cloacal es la zona equivalente en el extremo caudal del disco. Marca el futuro extremo posterior del tubo digestivo. Al romperse, formará las aberturas anorrectales y urogenitales.
Las membranas bucofaríngea y cloacal son preguntas clásicas de examen. Son las únicas regiones del disco trilaminar sin mesodermo intraembrionario. La membrana bucofaríngea marca el límite entre el ectodermo oral y el endodermo digestivo: su posición explica por qué el esmalte dental (derivado del ectodermo) y el epitelio del resto de la boca tienen orígenes distintos.
11.5. Formación de la notocorda
A través del nódulo de Hensen, las células epiblásticas migran en dirección craneal por la línea media, interponiéndose entre el ectodermo y el endodermo para formar un cordón celular compacto: el proceso notocordal. Este proceso se extiende desde el nódulo hasta la membrana bucofaríngea.
El proceso notocordal se transforma progresivamente en la notocorda, una estructura cilíndrica sólida que ocupa la línea media del embrión. La transformación implica la fusión transitoria del proceso notocordal con el endodermo subyacente y la posterior separación. Durante este proceso se forma brevemente el canal neuroentérico, una comunicación entre la cavidad amniótica y el saco vitelino a través de la notocorda.
La notocorda es una estructura mesenquimal cilíndrica y transitoria que ocupa la línea media del embrión entre la membrana bucofaríngea y el nódulo de Hensen. Es el principal organizador del desarrollo axial: induce al ectodermo suprayacente a diferenciarse en placa neural (inicio de la neurulación) y proporciona señales de posición que organizan el mesodermo paraxial en somitas.
La notocorda tiene dos funciones principales durante el desarrollo:
- Función inductora: la notocorda secreta factores de señalización (principalmente Sonic Hedgehog, SHH) que inducen al ectodermo que la recubre a engrosarse y formar la placa neural, iniciando la neurulación.
- Función mecánica: actúa como un eje rígido provisional que mantiene la forma del embrión durante el plegamiento.
En el adulto, la notocorda prácticamente desaparece. Su único remanente es el núcleo pulposo de los discos intervertebrales, la región gelatinosa central del disco que actúa como amortiguador entre las vértebras.
El cordoma es un tumor maligno raro que deriva de restos notocordales persistentes. Se presenta típicamente en los extremos del eje axial: el 50% en la región sacrococcígea y el 35% en la región esfenooccipital (clivus). Su comportamiento es localmente agresivo y tiene tendencia a la recidiva local tras la cirugía. El diagnóstico histológico se basa en la identificación de células fisalíforas (células vacuoladas características) y la positividad inmunohistoquímica para brachyury, un factor de transcripción específico de la notocorda.
11.6. Organización del mesodermo intraembrionario
El mesodermo intraembrionario formado durante la gastrulación se organiza en tres regiones con destinos distintos a lo largo del eje medio-lateral.
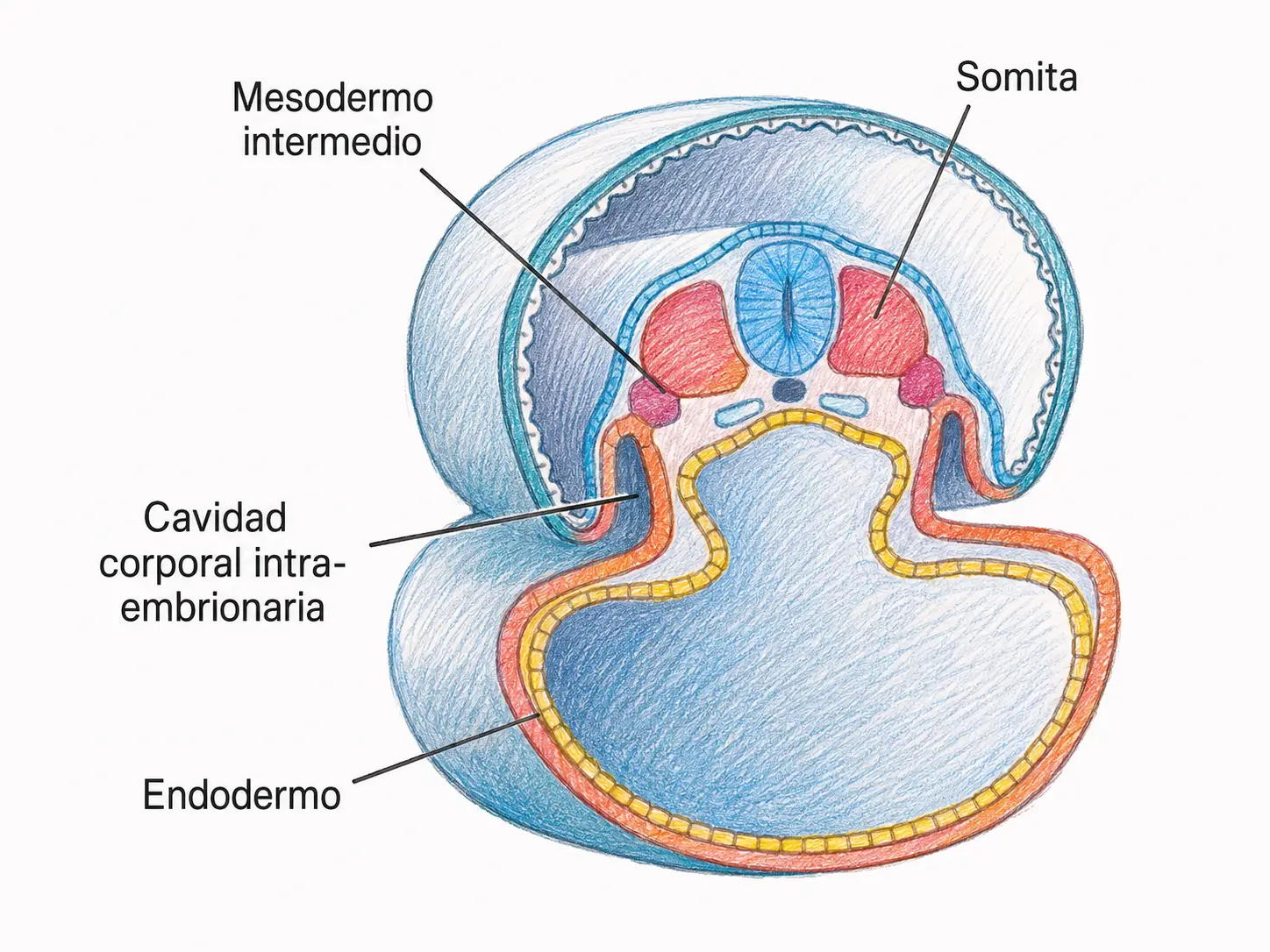
- El mesodermo paraxial es la región más medial, flanqueando a la notocorda a ambos lados. A partir del día 20, el mesodermo paraxial se segmenta en bloques compactos denominados somitas, que aparecen en dirección céfalo-caudal a un ritmo de 3-4 pares por día. Al final del periodo embrionario habrá entre 42 y 44 pares de somitas. Cada somita se subdivide posteriormente en tres regiones:
- El esclerotomo (que formará las vértebras y las costillas).
- El miotomo (que formará la musculatura esquelética).
- El dermotomo (que formará la dermis).
- El desarrollo de los somitas se estudia en detalle en el T12 (→ T12: somitas y formación de las extremidades).
- El mesodermo intermedio es la región de transición entre el mesodermo paraxial y el lateral. Da lugar al sistema urogenital: los túbulos del pronefros, el mesonefros y el metanefros, así como las gónadas (→ T22: desarrollo del aparato urogenital).
- El mesodermo lateral es la región más periférica. Se divide en dos hojas por la aparición del celoma intraembrionario (la futura cavidad corporal):
- La hoja somatopleural (adyacente al ectodermo), que formará la pared del cuerpo y las extremidades.
- La hoja esplacnopleural (adyacente al endodermo), que formará las paredes del aparato digestivo y respiratorio y el aparato cardiovascular.
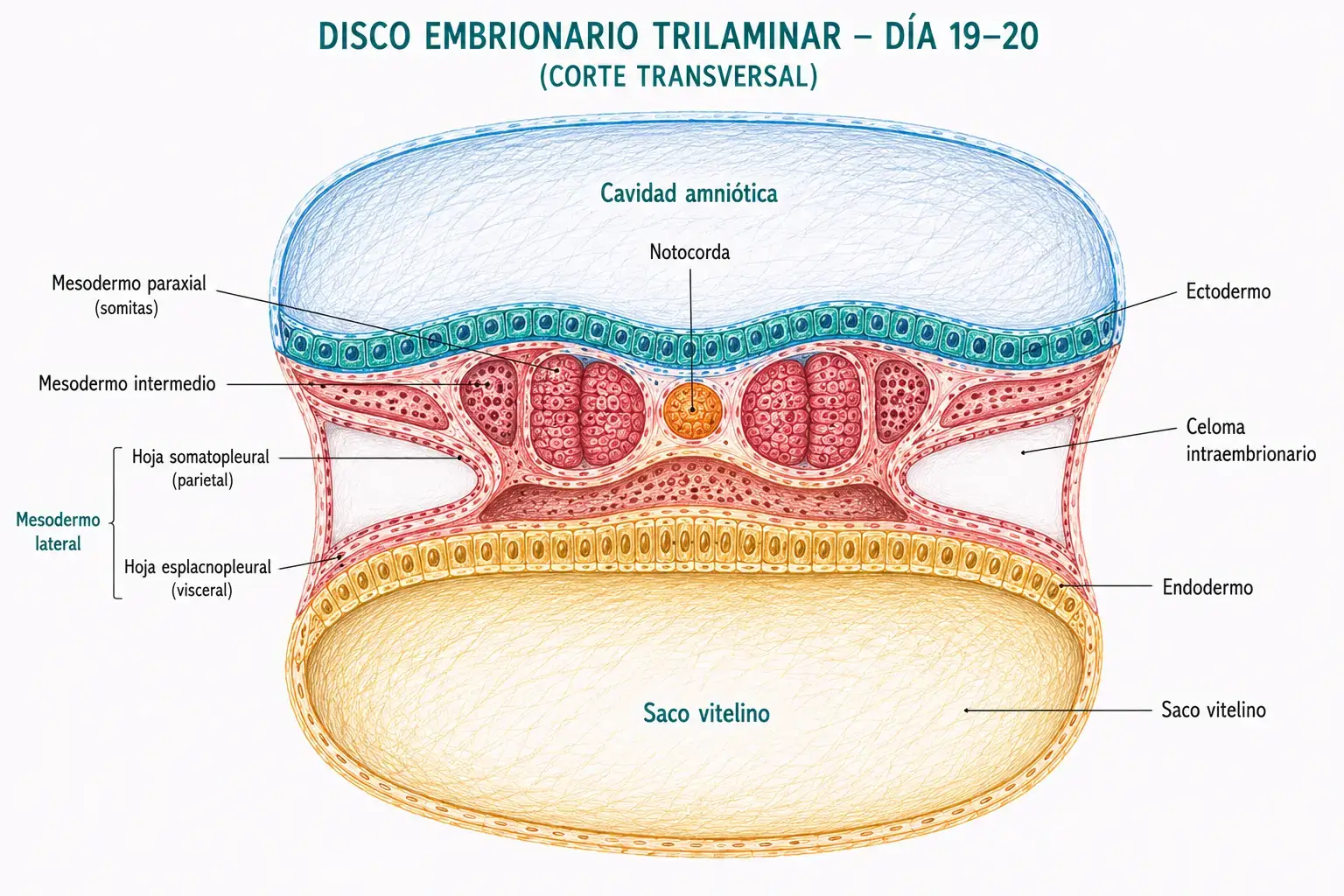
La segmentación del mesodermo en paraxial, intermedio y lateral no es arbitraria: refleja un gradiente de señalización a lo largo del eje medio-lateral establecido durante la gastrulación. Cuanto más medial es el mesodermo, más influenciado está por las señales de la notocorda (SHH) y más dorsal será su destino (vértebras, musculatura axial). Cuanto más lateral, más influenciado por las señales del ectodermo lateral y más ventral será su destino (cavidades, corazón, extremidades).
11.7. El celoma intraembrionario
Hacia el final de la tercera semana, dentro del mesodermo lateral aparecen pequeñas cavidades que confluyen formando el celoma intraembrionario, la futura cavidad corporal del embrión. Esta cavidad se comunica transitoriamente con el celoma extraembrionario (la cavidad coriónica) a través de los bordes laterales del disco embrionario.
El celoma intraembrionario se dividirá posteriormente en tres cavidades:
- La cavidad pericárdica (que alojará el corazón).
- Las dos cavidades pleurales (que alojarán los pulmones).
- La cavidad peritoneal (que alojará las vísceras abdominales).
La separación de estas cavidades ocurre durante las semanas 4 a 8.
11.8. Establecimiento de los ejes corporales
La gastrulación también establece los tres ejes espaciales del organismo.
- El eje cráneo-caudal (o antero-posterior) queda definido por la orientación de la línea primitiva y la dirección de migración de las células que forman la notocorda. El polo craneal es el extremo donde se formará la membrana bucofaríngea y donde la notocorda tiene su extremo anterior.
- El eje dorso-ventral queda definido por la posición relativa de las hojas: el ectodermo es dorsal (cubre la superficie del embrión), el endodermo es ventral (tapiza el futuro tubo digestivo) y el mesodermo es intermedio.
- El eje izquierda-derecha es el más complejo. Su establecimiento depende de la actividad ciliar de las células del nódulo de Hensen, que generan una corriente de líquido (flujo nodal) dirigida hacia la izquierda. Esta corriente arrastra morfógenos como Nodal y activa en el lado izquierdo una cascada de señalización que determinará la asimetría visceral (situs): el corazón a la izquierda, el hígado a la derecha, el estómago a la izquierda, etc.
El situs inversus es la condición en la que todos los órganos internos están en posición especular a la normal. Ocurre cuando el flujo nodal falla o está ausente, como en el síndrome de Kartagener (discinesia ciliar primaria), en el que los cilios del nódulo de Hensen son inmóviles. En ausencia de la señal direccional izquierda, la lateralización de los órganos queda al azar (50% normal, 50% invertida). El situs inversus totalis aislado no produce síntomas, pero el situs ambiguus (heterotaxia), en el que algunos órganos están en posición normal y otros invertida, se asocia frecuentemente a cardiopatías congénitas complejas.
11.9. Fin de la tercera semana: el disco trilaminar
Al final de la tercera semana, la situación del embrión es la siguiente:
- El disco bilaminar se ha transformado en un disco trilaminar con las tres hojas germinativas establecidas.
- La notocorda ocupa la línea media.
- Han aparecido los primeros somitas (3-4 pares en el día 20).
- El celoma intraembrionario ha comenzado a formarse.
- El corazón primitivo está iniciando su desarrollo en el mesodermo cardiogénico, que es la región del mesodermo lateral situado por delante de la membrana bucofaríngea.
- La placa neural ha comenzado a formarse en el ectodermo suprayacente a la notocorda, señalando el inicio de la neurulación.
Al final de la tercera semana el embrión mide apenas 1,5-2 mm. Sin embargo, en estos siete días ha pasado de ser un disco de dos capas indiferenciadas a tener las tres hojas germinativas, los ejes corporales establecidos, los primeros somitas formados y los esbozos del sistema nervioso y del corazón iniciados. La tercera semana es, en proporción, el periodo de mayor complejidad morfogenética de todo el desarrollo humano.
11.10. Plegamiento del embrión: del disco al cilindro
Al final de la tercera semana y durante la cuarta, el disco trilaminar plano se transforma en un embrión tridimensional de forma cilíndrica mediante el plegamiento corporal. Este proceso es la consecuencia mecánica del crecimiento diferencial entre las distintas regiones del disco. El embrión crece mucho más rápido en su eje longitudinal y en su región cefálica que en sus bordes laterales, lo que genera fuerzas que doblan el disco sobre sí mismo.
El plegamiento ocurre simultáneamente en tres direcciones y es irreversible: una vez iniciado, establece la forma corporal básica que se mantendrá durante todo el desarrollo posterior.
11.10.1. Plegamiento cefálico
El extremo craneal del disco, donde la placa neural y el mesodermo cardiogénico crecen con especial intensidad, se dobla ventralmente. Como consecuencia, la membrana bucofaríngea (que inicialmente miraba hacia craneal) queda orientada hacia ventral y caudal, en la posición del futuro estomodeo. El mesodermo cardiogénico, que estaba cranealmente a la membrana bucofaríngea, queda arrastrado hacia la posición ventral del embrión, donde formará el corazón en la cavidad pericárdica.
El septum transversum, inicialmente situado cranealmente al corazón, queda desplazado caudalmente hasta su posición definitiva entre el corazón y el hígado en desarrollo, donde formará el centro tendinoso del diafragma.
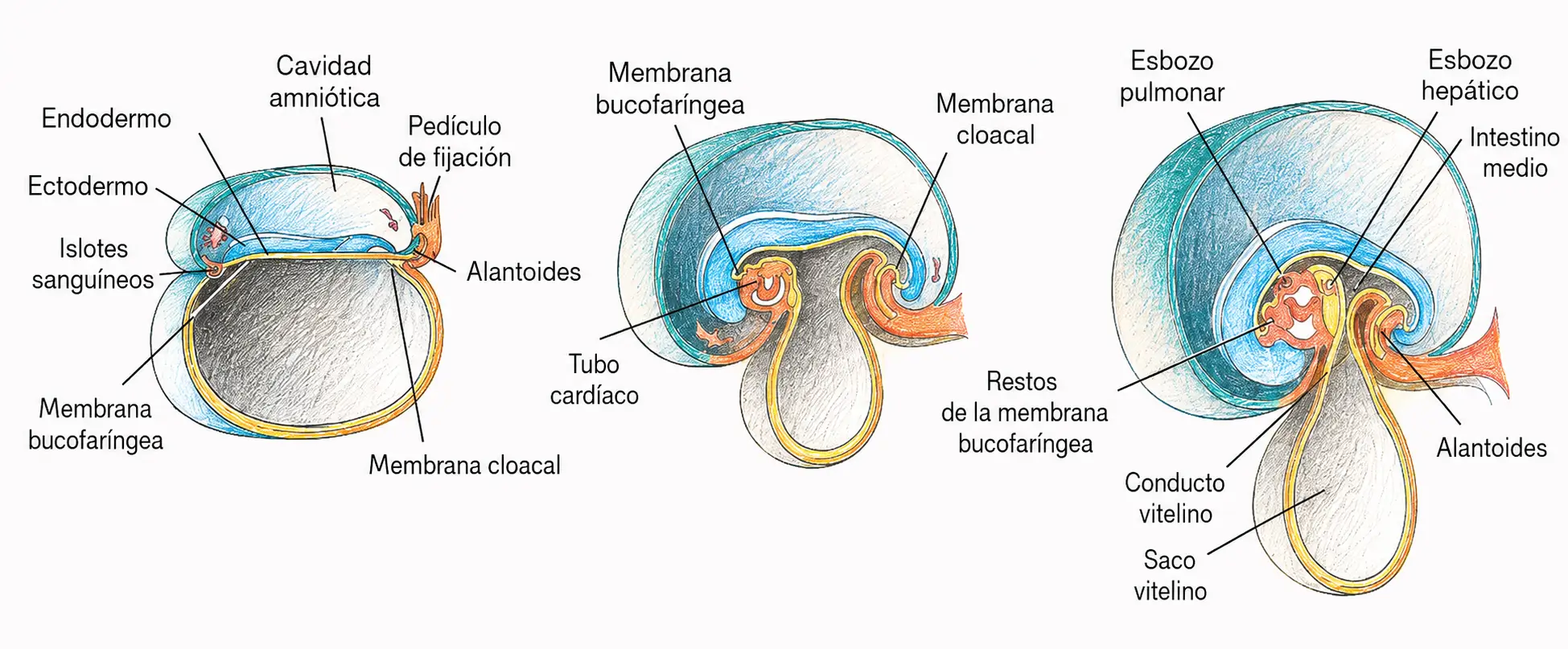
El plegamiento cefálico explica una aparente paradoja anatómica: el mesodermo cardiogénico estaba craneal a la membrana bucofaríngea antes del plegamiento, pero el corazón adulto está caudal a la boca. El plegamiento cefálico invierte esta relación. Sin conocer el plegamiento, la posición del corazón respecto a la faringe en el adulto resulta incomprensible.
11.10.2. Plegamiento lateral
Los bordes laterales del disco se doblan ventralmente a ambos lados, aproximando las dos mitades del embrión hacia la línea ventral. Este movimiento tiene consecuencias múltiples y simultáneas.
Los dos tubos cardiacos endocárdicos, situados uno a cada lado de la línea media, son aproximados por el plegamiento lateral y se fusionan formando el tubo cardiaco único. El endodermo, que tapizaba la cara ventral del disco, queda incorporado al interior del embrión formando el intestino primitivo: un tubo endodérmico cerrado cranealmente (intestino anterior) y caudalmente (intestino posterior), con comunicación temporal con el saco vitelino a través del conducto vitelino en la región media (intestino medio).
El celoma intraembrionario, que era una cavidad abierta en el disco, queda encerrado dentro del cuerpo del embrión formando la cavidad corporal primitiva, que se comunicará transitoriamente con el celoma extraembrionario antes de cerrarse.
La somatopleura (ectodermo + mesodermo somatopleural lateral) forma la pared del cuerpo del embrión. La esplacnopleura (endodermo + mesodermo esplacnopleural) rodea el intestino primitivo.
11.10.3. Plegamiento caudal
El extremo caudal del disco se dobla ventralmente, incorporando la alantoides y parte del saco vitelino al interior del embrión. La membrana cloacal queda orientada ventralmente. El pedículo de fijación, que conectaba el embrión con el trofoblasto, queda comprimido junto con el conducto vitelino formando el cordón umbilical primitivo.
11.10.4. Resultado del plegamiento: el embrión cilíndrico
Al completarse el plegamiento, hacia el final de la cuarta semana, el embrión ha pasado de ser un disco plano trilaminar a ser un cilindro con:
Una pared corporal cerrada (somatopleura) que delimita el interior del embrión del exterior. Un intestino primitivo endodérmico en posición central, cerrado en sus extremos por las membranas bucofaríngea y cloacal. Una cavidad celómica intraembrionaria que rodea las vísceras. Un corazón en posición ventral dentro de la cavidad pericárdica. Un cordón umbilical que conecta el embrión con la placenta.
El plegamiento explica la posición de múltiples estructuras adultas que de otro modo resultarían inexplicables: el corazón ventral (mesodermo cardiogénico era craneal), el ombligo (punto de cierre del plegamiento lateral donde queda el conducto vitelino y el pedículo de fijación), la posición del diafragma (septum transversum desplazado caudalmente por el plegamiento cefálico) y la orientación de la membrana cloacal (desplazada ventralmente por el plegamiento caudal). Estas cuatro preguntas tienen la misma respuesta: el plegamiento embrionario.
El plegamiento marca el inicio del periodo embrionario propiamente dicho (semanas 4 a 8), durante el cual se forman todos los órganos mediante organogénesis. A partir de este momento, el embrión tiene la arquitectura corporal básica establecida y cada sistema orgánico comienza su desarrollo específico a partir de las hojas germinativas en sus posiciones correctas.
El plegamiento cefálico explica la posición del corazón y es el punto de partida de la cardiogénesis (ver Tema 14 · Cardiogénesis).
El plegamiento lateral explica la formación del intestino primitivo, punto de partida del aparato digestivo (Tema 22 · Aparato digestivo) y del Tema 21 · Aparato respiratorio.
El plegamiento caudal explica la formación de la cloaca y es el punto de partida del aparato urogenital y del polo caudal (Tema 23 · Aparato urinario y Tema 24 · Aparato genital y polo caudal).
