8.1. El encuentro entre gametos
Para que la fecundación pueda ocurrir, el espermatozoide debe recorrer el tracto genital femenino hasta alcanzar el ovocito en la trompa de Falopio. Este recorrido es largo y adverso: de los cientos de millones de espermatozoides del eyaculado, solo unos pocos cientos llegan a la ampolla tubárica.
Tras la eyaculación en la vagina, los espermatozoides deben atravesar el moco cervical, ascender por el útero y entrar en las trompas de Falopio. El transporte se facilita por las contracciones uterinas y el movimiento ciliar del epitelio tubárico. La mayor parte de los espermatozoides quedan retenidos o destruidos a lo largo del camino: en el moco cervical, en la unión útero-tubárica y en el istmo de la trompa. Solo en el momento periovulatorio, cuando los estrógenos han hecho el moco cervical más fluido y permeable, el paso es posible.
Los espermatozoides conservan su capacidad fecundante durante unas 24-48 horas en el tracto genital femenino. El ovocito secundario, por su parte, permanece fecundable durante unas 12-24 horas tras la ovulación. La ventana fértil del ciclo menstrual es, por tanto, de unos 5-6 días (considerando la supervivencia espermática preovulatoria).
La determinación de la ventana fértil tiene importancia tanto para buscar el embarazo como para evitarlo mediante métodos naturales. Los métodos basados en el conocimiento de la fertilidad (como el método Billings o los kits de detección de LH urinario) se basan en identificar el momento de la ovulación para localizar y actuar en consecuencia sobre esta ventana de 5-6 días.
Durante el ascenso por el tracto femenino, los espermatozoides completan la capacitación si no lo habían hecho antes. La capacitación modifica la membrana plasmática del espermatozoide, activa canales de calcio y prepara al espermatozoide para la hiperactivación y la reacción acrosómica (ver Tema 2 · Espermatogénesis).
8.2. Penetración de la corona radiada
El ovocito secundario ovulado llega a la ampolla tubárica rodeado por dos capas protectoras:
- La zona pelúcida (capa de glucoproteínas).
- Externa a la zona pelúcida está la corona radiada, formada por las células del cúmulo oóforo que han acompañado al ovocito desde el folículo.
El espermatozoide hiperactivado penetra la corona radiada mediante dos mecanismos combinados.
- La hialuronidasa presente en la superficie del acrosoma digiere el ácido hialurónico de la matriz extracelular que une las células de la corona.
- La potente motilidad flagelar del espermatozoide capacitado le permite abrirse paso mecánicamente entre las células foliculares. La penetración de la corona radiada no requiere todavía la reacción acrosómica.
8.3. Reacción acrosómica y penetración de la zona pelúcida
La zona pelúcida es una barrera glucoproteica densa que rodea el ovocito. Está compuesta principalmente por tres glucoproteínas: ZP1 (estructural), ZP2 (receptora tras la fecundación) y ZP3 (receptora primaria del espermatozoide).
Cuando el espermatozoide contacta con la ZP3, esta glucoproteína actúa como ligando sobre receptores de la membrana plasmática del espermatozoide, desencadenando la reacción acrosómica.
La reacción acrosómica es la exocitosis del contenido del acrosoma, desencadenada por el contacto del espermatozoide con la ZP3 de la zona pelúcida. Implica la fusión de la membrana externa del acrosoma con la membrana plasmática del espermatozoide y la liberación al espacio extracelular de las enzimas hidrolíticas acrosómicas (principalmente acrosina e hialuronidasas). Estas enzimas digieren localmente la zona pelúcida, creando un canal por el que el espermatozoide penetra.
En la reacción acrosómica, la hiperactivación flagelar del espermatozoide capacitado, combinada con la actividad enzimática, ejerce una fuerza mecánica que permite atravesar la zona pelúcida. El espermatozoide que ha completado la reacción acrosómica queda con la membrana interna del acrosoma expuesta, que será la que participe en la fusión con la membrana del ovocito.
La reacción acrosómica debe producirse en la zona pelúcida, no antes. Un espermatozoide que experimenta la reacción acrosómica prematuramente (antes de alcanzar el ovocito) pierde su capacidad fecundante porque ya no dispone de las enzimas necesarias para atravesar la zona pelúcida. La capacitación regula este proceso para que la reacción acrosómica se desencadene en el lugar correcto.
8.4. Fusión de membranas y activación del ovocito
Tras atravesar la zona pelúcida, el espermatozoide alcanza el espacio perivitelino y entra en contacto con la membrana plasmática del ovocito secundario. La fusión de ambas membranas está mediada por proteínas específicas: en el espermatozoide, la proteína IZUMO1; en el ovocito, la proteína JUNO. La interacción IZUMO1-JUNO es el paso molecular esencial de la fusión de membranas.
Tras la fusión, el contenido del espermatozoide (núcleo y centríolo principalmente) entra en el citoplasma del ovocito. La cola del espermatozoide permanece fuera o es degradada.
La entrada del espermatozoide desencadena la activación del ovocito, una cascada de señalización intracelular iniciada por la liberación de calcio desde el retículo endoplásmico del ovocito. Esta liberación de calcio ocurre en forma de ondas sucesivas que se propagan por todo el citoplasma del ovocito.
La liberación de calcio intracelular en el ovocito tras la fecundación es el disparador de todos los eventos de activación: resolución del segundo arresto meiótico, exocitosis de los gránulos corticales y reanudación de las divisiones de segmentación. El factor espermático responsable de desencadenar esta liberación de calcio se ha identificado como la fosfolipasa C zeta (PLCζ), introducida por el espermatozoide en el citoplasma del ovocito.
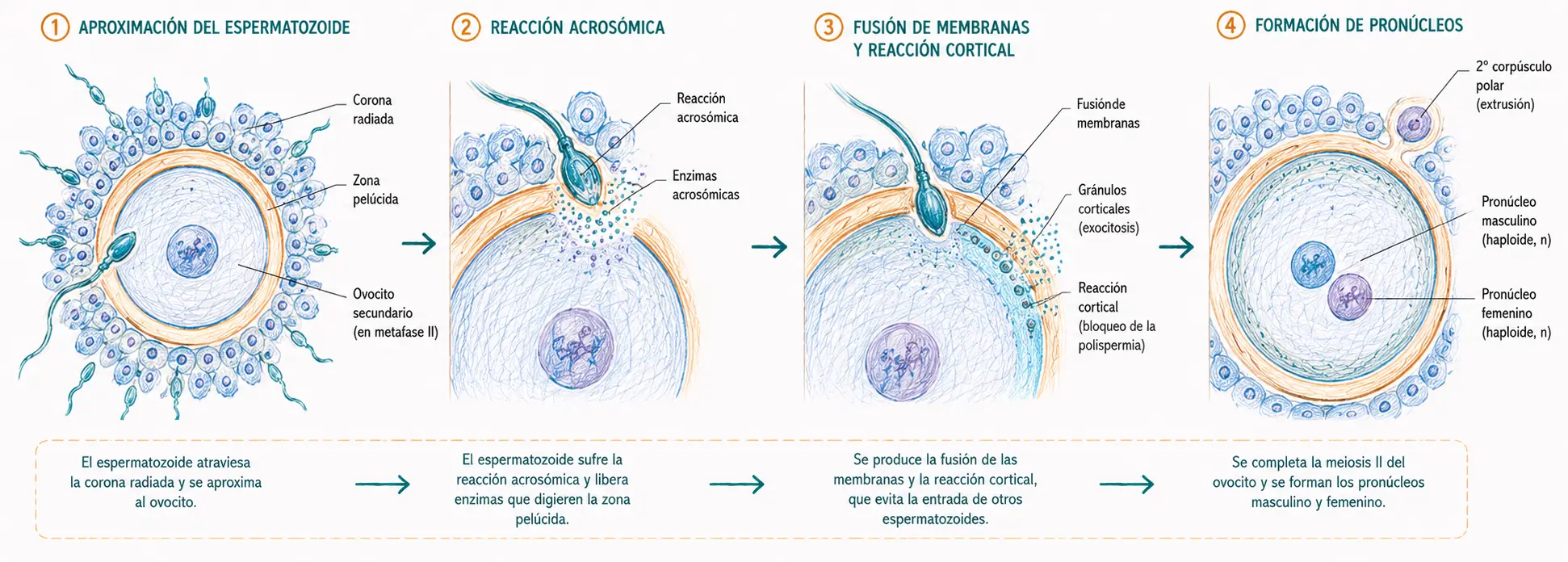
8.5. Prevención de la polispermia
La polispermia, es decir, la entrada de más de un espermatozoide en el ovocito, es incompatible con el desarrollo normal: produciría un cigoto triploide o poliploide inviable. Existen dos mecanismos sucesivos que la previenen.
- Reacción rápida (bloqueo eléctrico): en los segundos inmediatamente posteriores a la fusión de membranas, el potencial de membrana del ovocito cambia bruscamente de negativo a positivo (despolarización). Este cambio en el potencial eléctrico impide físicamente la fusión de nuevos espermatozoides con la membrana ovocitaria. Es un mecanismo rápido pero transitorio.
- Reacción lenta (reacción cortical): en los minutos siguientes, las ondas de calcio desencadenan la exocitosis de los gránulos corticales, vesículas secretoras situadas justo debajo de la membrana plasmática del ovocito. El contenido de los gránulos corticales (enzimas como la ovoperoxidasa y proteínas estructurales) es liberado al espacio perivitelino y modifica la zona pelúcida: endurece su estructura y destruye los receptores ZP3. Esta modificación se denomina reacción de zona y constituye un bloqueo permanente a la polispermia.
La prevención de la polispermia tiene dos fases: (1) bloqueo rápido, por despolarización de membrana, que actúa en segundos; (2) bloqueo lento o reacción cortical, por exocitosis de gránulos corticales que modifican la zona pelúcida, que actúa en minutos. En el examen puede preguntarse cuál de los dos es reversible (el rápido) y cuál permanente (el lento).
La polispermia es una de las causas de fracaso en la fecundación in vitro convencional, en la que varios espermatozoides capacitados pueden contactar simultáneamente con el ovocito en el medio de cultivo. La inyección intracitoplasmática de espermatozoide (ICSI) elimina este problema al introducir mecánicamente un único espermatozoide en el citoplasma del ovocito.
8.6. Formación del cigoto
Tras la activación del ovocito, la meiosis II se completa:
- El ovocito secundario se divide asimétricamente produciendo el ovocito maduro y el segundo corpúsculo polar. El núcleo del ovocito maduro se denomina pronúcleo femenino (n = 23).
- Paralelamente, el núcleo del espermatozoide se descondensa: las protaminas son sustituidas por histonas del ovocito y se forma el pronúcleo masculino (n = 23).
Ambos pronúcleos migran hacia el centro del ovocito y se aproximan. En el humano, los pronúcleos no llegan a fusionarse antes de la primera división de segmentación: sus membranas nucleares se disuelven, los cromosomas de ambos pronúcleos se condensan en la misma placa metafásica y la primera división de segmentación distribuye los cromosomas entre las dos células hijas. Este proceso de unión de los materiales genéticos de ambos pronúcleos se denomina anfimixis y marca formalmente el inicio del desarrollo embrionario.
El cigoto es la célula diploide (2n = 46) resultante de la fecundación, desde el momento en que los pronúcleos se aproximan hasta la primera división de segmentación. Es la primera célula del nuevo individuo.
Los resultados biológicos de la fecundación son cuatro. Primero, la restauración de la ploídía diploide (2n = 46 cromosomas), combinando 23 cromosomas de origen materno y 23 de origen paterno. Segundo, la determinación del sexo cromosómico: si el espermatozoide porta cromosoma Y el cigoto será 46,XY (varón); si porta X será 46,XX (mujer). Tercero, el inicio de las divisiones de segmentación, que transformarán el cigoto en el embrión multicelular. Cuarto, la activación del genoma embrionario, que ocurre hacia el estadio de 8 células (en torno al día 3 de desarrollo).
8.7. Embarazo ectópico
En condiciones normales, el cigoto es transportado por el epitelio ciliado y las contracciones de la trompa hacia la cavidad uterina, mientras realiza sus primeras divisiones de segmentación. Ocasionalmente, el embrión se implanta fuera del útero: el caso más frecuente es la implantación en la propia trompa de Falopio, produciendo un embarazo ectópico tubárico.
El embarazo ectópico tubárico es una urgencia médica. La trompa no puede acomodar el crecimiento embrionario y, si no se trata, se rompe produciendo una hemorragia intraabdominal grave. Los factores de riesgo incluyen antecedentes de enfermedad inflamatoria pélvica, cirugía tubárica previa y tabaquismo, todos ellos relacionados con alteraciones de la motilidad tubárica. El diagnóstico precoz se realiza mediante ecografía transvaginal y determinación de la beta-hCG sérica.
