12.1. Inducción de la placa neural
La neurulación comienza con un acto de inducción: la notocorda, recién formada en la línea media del mesodermo, secreta factores de señalización que modifican el destino del ectodermo que la recubre. El principal factor inductor es SHH (Sonic Hedgehog), aunque también participan BMP antagonistas (Noggin, Chordin, Folistatin) secretados por la notocorda y por el organizador del nódulo de Hensen.
Estos factores inhiben la vía BMP en el ectodermo suprayacente. En ausencia de señalización BMP, las células ectodérmicas activan un programa de diferenciación neural: se elongan, adquieren morfología columnar y expresan marcadores neurales como Sox2 y N-caderina. El resultado es el neuroectodermo o placa neural: una zona engrosada y aplanada de ectodermo que ocupa la línea media dorsal del embrión, por encima de la notocorda.
La inducción neural es un ejemplo clásico del principio de inducción embrionaria: un tejido (la notocorda) modifica el destino de un tejido vecino (el ectodermo) mediante señales moleculares difusibles. Sin la presencia de la notocorda en posición correcta, el ectodermo suprayacente no forma placa neural y el sistema nervioso central no se desarrolla.
La placa neural es inicialmente más ancha en su extremo craneal, que dará lugar al encéfalo, y más estrecha en su extremo caudal, que dará lugar a la médula espinal. Esta diferencia de anchura ya refleja el mayor volumen que tendrá el encéfalo respecto a la médula en el adulto.
12.2. Neurulación primaria: formación y cierre del tubo neural
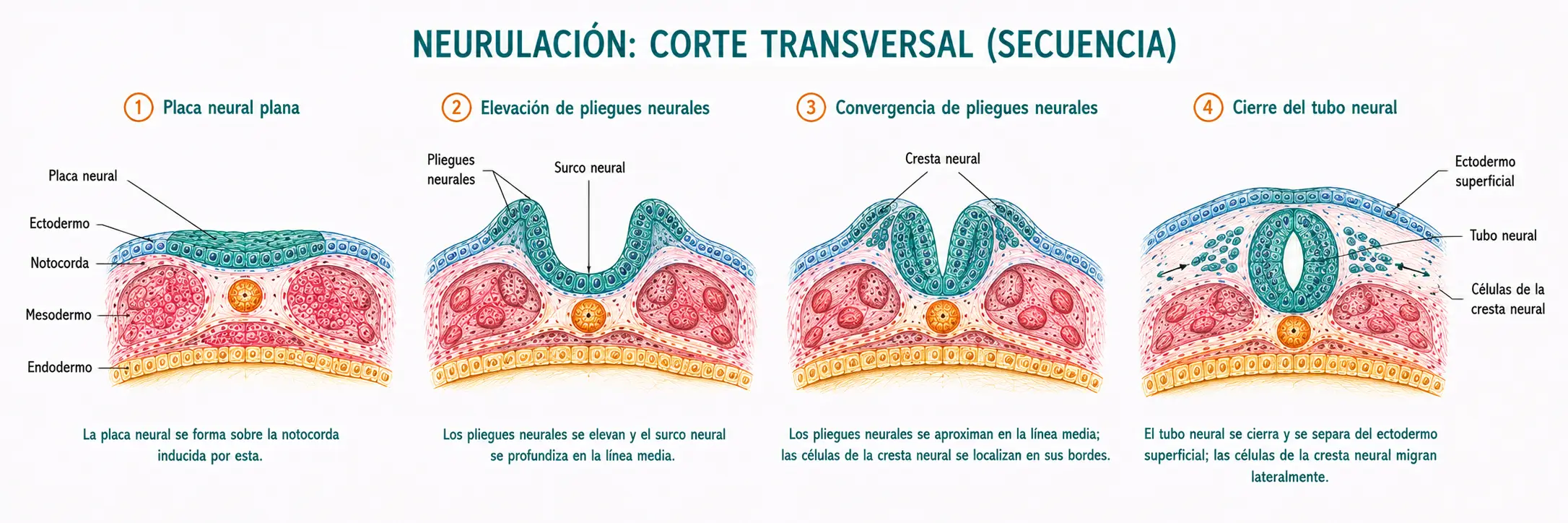
La neurulación primaria comprende la secuencia de plegamiento de la placa neural hasta formar el tubo neural cerrado.
12.2.1. Plegamiento de la placa neural
Los bordes laterales de la placa neural se elevan formando los pliegues neurales, mientras que la región central se deprime formando el surco neural. Los pliegues neurales crecen hacia la línea media y comienzan a aproximarse dorsalmente.
El plegamiento está impulsado por cambios en la forma de las células neuroepiteliales: las células de las zonas de bisagra mediales y laterales de la placa neural adquieren morfología en cuña (más estrechas en su polo apical), lo que curva la lámina. Este cambio de forma está mediado por el citoesqueleto de actina y miosina.
12.2.2. Cierre del tubo neural
Los pliegues neurales se aproximan en la línea media dorsal y se fusionan, formando el tubo neural. La fusión comienza aproximadamente a nivel del quinto somita, hacia el día 22, y progresa bidireccionalmente (hacia craneal y hacia caudal) de forma simultánea.
Los extremos del tubo que todavía no se han cerrado se denominan neuroporos.
- El neuroporo craneal (anterior) se cierra hacia el día 25.
- El neuroporo caudal (posterior) se cierra hacia el día 27-28.
Con el cierre de ambos neuroporos, el tubo neural queda completamente sellado y separado del ectodermo de superficie, que se cierra por encima de él.
El cierre bidireccional del tubo neural explica el patrón de los defectos del tubo neural: el fallo en el cierre del neuroporo craneal produce anencefalia (ausencia de bóveda craneal y de gran parte del encéfalo), incompatible con la vida. El fallo en el cierre del neuroporo caudal produce espina bífida a nivel lumbosacro, con consecuencias variables según la extensión de la lesión.
Los defectos del tubo neural (DTN) tienen una incidencia de aproximadamente 1-2 por 1.000 nacidos vivos y son las malformaciones congénitas del sistema nervioso más frecuentes. El ácido fólico es cofactor de enzimas implicadas en la síntesis de purinas y timidina, esenciales para la proliferación celular acelerada del tubo neural. La suplementación periconcepcional con 0,4-4 mg/día de ácido fólico reduce la incidencia de DTN en un 50-70%. Por este motivo, la suplementación se recomienda a todas las mujeres en edad fértil desde al menos un mes antes de la concepción y durante el primer trimestre.
12.2.3. Neurulación secundaria
La porción más caudal del tubo neural (región sacra inferior y coccígea) no se forma por plegamiento de la placa neural, sino por un proceso diferente denominado neurulación secundaria: las células mesenquimatosas de la masa caudal del embrión forman un cordón celular sólido que posteriormente se canaliza y se fusiona con el tubo neural formado por neurulación primaria.
12.3. Histogénesis del tubo neural
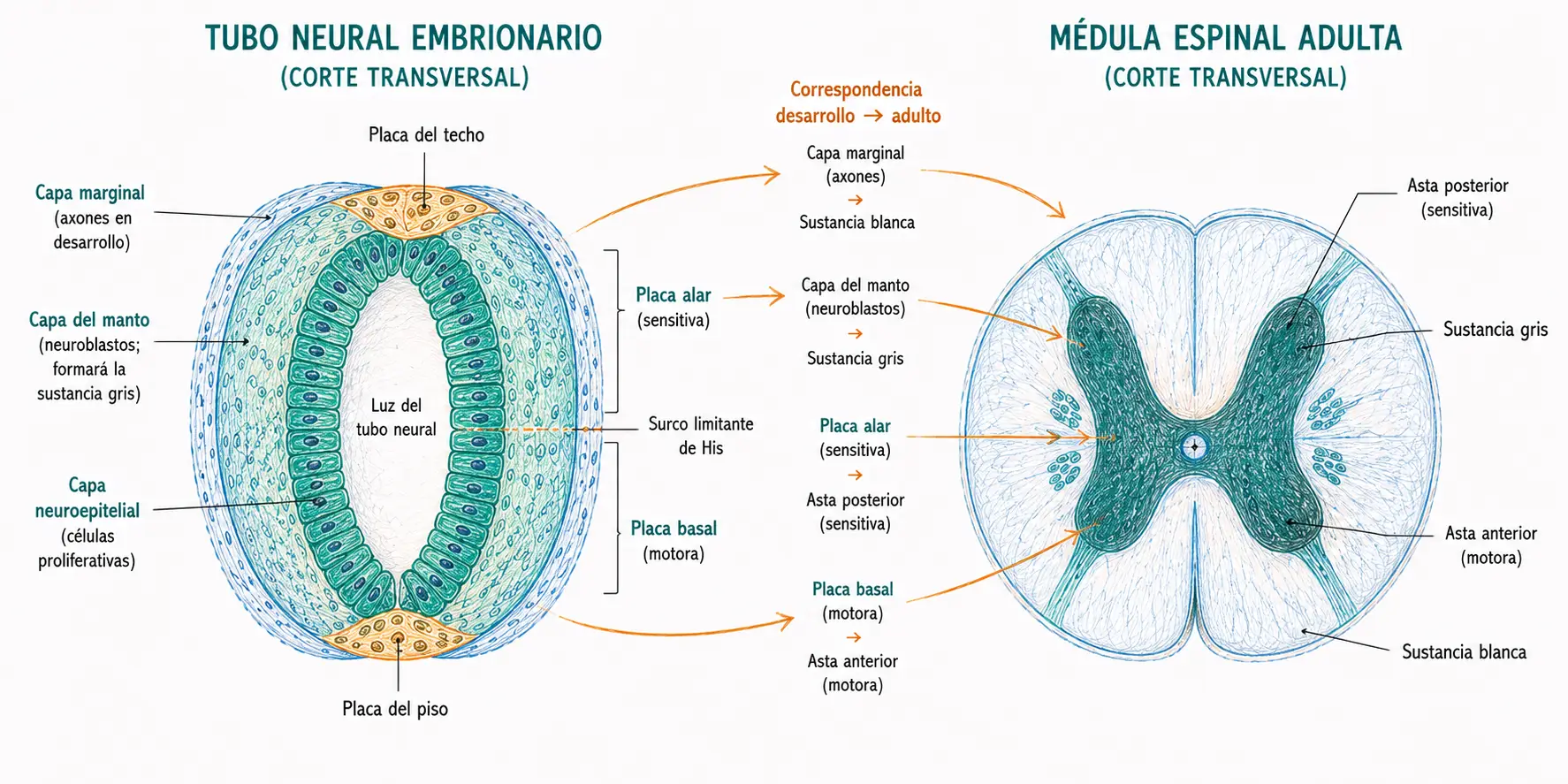
Una vez cerrado, la pared del tubo neural prolifera activamente y se organiza en tres capas concéntricas desde la luz hacia el exterior.
- La capa neuroepitelial (o zona ventricular) es la capa más interna, en contacto con la luz del tubo. Está formada por células neuroepiteliales en intensa proliferación mitótica. Es la capa generadora: produce todos los neuroblastos y glioblastos del sistema nervioso central. Las mitosis ocurren en la superficie que da a la luz del tubo.
- La capa del manto (o zona intermedia) se forma cuando los neuroblastos postmitóticos migran desde la capa neuroepitelial hacia el exterior. Los neuroblastos son células que han salido del ciclo celular y se están diferenciando en neuronas. Esta capa constituye la futura sustancia gris de la médula espinal, donde residirán los cuerpos de las neuronas.
- La capa marginal es la capa más externa. No contiene cuerpos neuronales sino los axones que emergen de los neuroblastos de la capa del manto. Con el tiempo estos axones se mielinizan, lo que da a esta capa un aspecto blanquecino: constituye la futura sustancia blanca de la médula espinal.
La correspondencia entre las capas embrionarias del tubo neural y las regiones adultas de la médula es pregunta frecuente:
- Capa neuroepitelial (proliferativa, desaparece en gran parte) → epéndimo del canal central.
- Capa del manto (cuerpos neuronales) → sustancia gris.
- Capa marginal (axones mielinizados) → sustancia blanca.
12.4. Placas basales, alares, del techo y del piso
La proliferación de neuroblastos en la capa del manto se produce de forma diferencial en las regiones ventral y dorsal del tubo, creando cuatro engrosamientos longitudinales.
- Los engrosamientos ventrales forman las placas basales, a ambos lados de la línea media ventral. Contienen neuronas motoras (eferentes) que inervarán la musculatura. En la médula adulta corresponden a las astas anteriores (ventrales), que contienen las motoneuronas alfa y gamma.
- Los engrosamientos dorsales forman las placas alares, a ambos lados de la línea media dorsal. Contienen neuronas sensitivas (aferentes) que reciben información del exterior. En la médula adulta corresponden a las astas posteriores (dorsales), que reciben las aferencias sensitivas de los ganglios de la raíz dorsal.
- El surco limitante de His es el surco longitudinal que separa la placa basal de la placa alar en cada lado del tubo. Marca la frontera entre el territorio motor (ventral) y el sensitivo (dorsal), que se conserva en la organización funcional de la médula adulta.
- Las regiones dorsales y ventrales de la línea media del tubo que no tienen neuroblastos forman respectivamente la placa del techo y la placa del piso. Estas placas actúan como organizadores moleculares que guían el crecimiento de los axones comisurales.
La organización dorso-ventral del tubo neural establecida durante la neurulación se conserva en la médula adulta: dorsal = sensitivo, ventral = motor. Esta regla, establecida por Sherrington a finales del siglo XIX como "ley de Bell-Magendie", tiene su explicación embriológica en la diferenciación de las placas alares y basales bajo la influencia de gradientes moleculares opuestos: SHH desde la notocorda y la placa del piso (que ventraliza el tubo, genera motoneuronas) y BMP desde la placa del techo (que dorsaliza el tubo, genera neuronas sensitivas).
Entre las placas basales y alares, en la región intermedia, se acumula una población neuronal adicional que formará el asta lateral o asta intermediolateral de la médula. Las neuronas de esta asta corresponden a las neuronas preganglionares del sistema nervioso autónomo simpático y solo están presentes a nivel torácico (T1-T12) y lumbar superior (L1-L2).
12.5. Células de la cresta neural
Las células de la cresta neural son una población celular que emerge de los bordes dorsolaterales del tubo neural en el momento de su cierre. Son exclusivas de los vertebrados y tienen dos propiedades excepcionales: capacidad de migración a larga distancia a través del embrión, y multipotencia, es decir, capacidad de diferenciarse en tipos celulares muy diversos según el microambiente que encuentran en su destino.
La cresta neural es una población de células neuroectodérmicas que sufren una transición epitelio-mesenquimal en el momento del cierre del tubo neural y migran extensamente por el embrión. Son a veces denominadas la "cuarta hoja germinativa" por la diversidad de sus derivados, comparable a la de las tres hojas clásicas.
12.5.1. Migración de las células de la cresta neural
Las células de la cresta neural utilizan dos rutas principales de migración.
- La ruta dorsolateral, entre el ectodermo y el somita, lleva a las células hacia la piel, donde se diferenciarán en melanocitos.
- La ruta ventral, a través de la mitad anterior de cada somita, lleva a las células hacia los ganglios sensitivos, los ganglios del sistema nervioso autónomo, la médula suprarrenal y otros destinos viscerales.
La migración está guiada por señales de atracción (efrinas, CXCL12) y de repulsión (semaforinas, efrinas B) que dirigen a las células a sus destinos correctos. Errores en esta guía producen malformaciones congénitas características.
12.5.2. Derivados de la cresta neural
Los derivados de la cresta neural son extraordinariamente diversos y conviene conocerlos agrupados por categorías:
- Sistema nervioso periférico: ganglios sensitivos de la raíz dorsal (ganglios espinales), ganglios sensitivos de los nervios craneales, ganglios del sistema nervioso autónomo (simpático y parasimpático), ganglios entéricos (plexos de Meissner y Auerbach).
- Sistema endocrino: células cromafines de la médula suprarrenal (que secretan adrenalina y noradrenalina), células paraganglionares, células C del tiroides (que secretan calcitonina).
- Piel: melanocitos de toda la superficie corporal.
- Esqueleto craneofacial: la mayor parte de los huesos y cartílagos de la cara y el cráneo (mandíbula, maxilar, huesos nasales, cartílagos laríngeos) derivan de la cresta neural craneal, no del mesodermo. Este origen explica por qué las malformaciones craneofaciales son frecuentes en síndromes que afectan a la migración de la cresta neural.
- Tejidos blandos craneofaciales: estroma de las glándulas salivares, cornea, tejido conectivo de los arcos faríngeos.
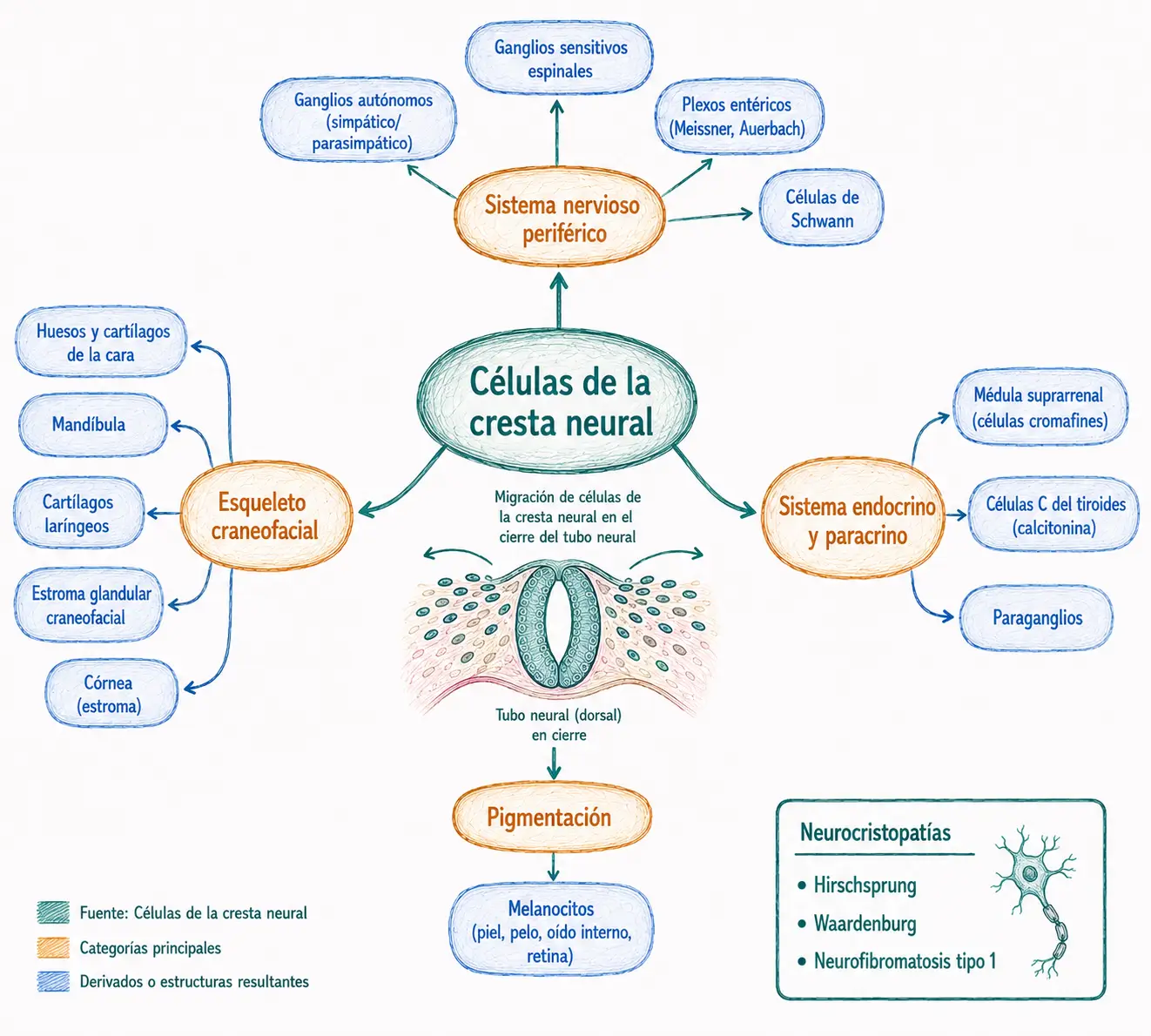
Las neurocristopatías son un grupo heterogéneo de enfermedades producidas por alteraciones en el desarrollo, la migración o la diferenciación de las células de la cresta neural. Incluyen:
- La enfermedad de Hirschsprung: ausencia de ganglios entéricos por fallo de migración al intestino, que produce megacolon congénito.
- El síndrome de Waardenburg: sordera y alteraciones de pigmentación por déficit de melanocitos y células del ganglio espiral de la cóclea.
- La neurofibromatosis tipo 1: tumores de células de Schwann y melanocitos.
- Otros síndromes craneofaciales.
La comprensión del origen en cresta neural de todos estos tejidos unifica aparentemente síndromes muy distintos bajo una misma patogenia.
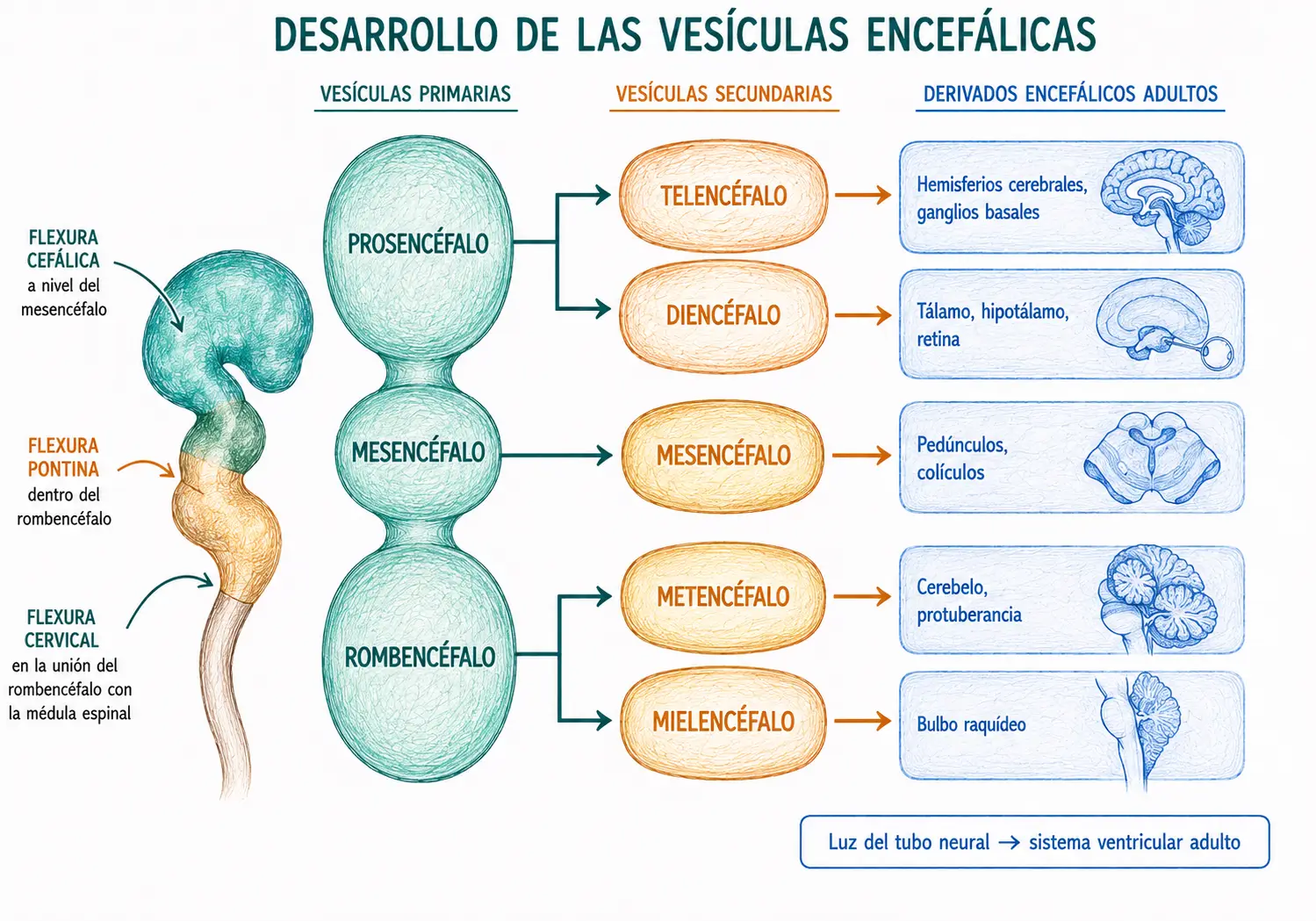
12.6. Vesículas encefálicas
La porción craneal del tubo neural crece mucho más rápidamente que la caudal y se dilata formando, hacia el final de la cuarta semana, tres vesículas encefálicas primarias:
- El prosencéfalo (vesícula anterior) da lugar a las estructuras del cerebro anterior.
- El mesencéfalo (vesícula media) da lugar al mesencéfalo adulto.
- El rombencéfalo (vesícula posterior) da lugar al cerebelo, la protuberancia y el bulbo raquídeo.
Hacia la quinta semana, las vesículas primarias se subdividen en cinco vesículas secundarias
- El prosencéfalo se divide en:
- Telencéfalo: formará los hemisferios cerebrales y los ganglios basales.
- Diencéfalo: formará el tálamo, el hipotálamo y la retina.
- El mesencéfalo no se subdivide.
- El rombencéfalo se divide en:
- Metencéfalo: formará el cerebelo y la protuberancia.
- Mielencéfalo: que formará el bulbo raquídeo.
| Vesícula primaria | Vesícula secundaria | Derivado adulto principal |
|---|---|---|
| Prosencéfalo | Telencéfalo | Hemisferios cerebrales, ganglios basales, hipocampo, amígdala |
| Diencéfalo | Tálamo, hipotálamo, epitálamo, retina, hipófisis posterior | |
| Mesencéfalo | Mesencéfalo | Pedúnculos cerebrales, colículos, sustancia negra |
| Rombencéfalo | Metencéfalo | Cerebelo, protuberancia (puente de Varolio) |
| Mielencéfalo | Bulbo raquídeo (médula oblongada) |
El tubo neural también experimenta tres flexuras durante su expansión craneal:
- La flexura cefálica: a nivel del mesencéfalo, hacia el día 24.
- La flexura cervical: a nivel de la unión rombencéfalo-médula.
- La flexura pontina: a nivel del metencéfalo, en dirección opuesta a las otras dos.
Estas flexuras pliegan el encéfalo en desarrollo y explican la geometría del encéfalo adulto.
12.7. El sistema ventricular y el líquido cefalorraquídeo
La luz del tubo neural persiste en el encéfalo adulto como el sistema ventricular. La luz del telencéfalo forma los ventrículos laterales (primero y segundo); la del diencéfalo forma el tercer ventrículo; la del mesencéfalo forma el acueducto de Silvio (acueducto cerebral); y la del rombencéfalo forma el cuarto ventrículo. La luz de la médula espinal persiste como el conducto ependimario (canal central).
El líquido cefalorraquídeo (LCR) se produce principalmente en los plexos coroideos de los ventrículos laterales, tercero y cuarto, y circula desde los ventrículos hacia el espacio subaracnoideo, donde es reabsorbido en las granulaciones aracnoideas. Su obstrucción produce hidrocefalia.
La hidrocefalia congénita puede producirse por estenosis del acueducto de Silvio (la causa más frecuente), por malformación de Chiari II (asociada al mielomeningocele, en la que el tronco del encéfalo y el cerebelo se hernian hacia el canal cervical obstruyendo la circulación del LCR) o por otras causas. El tratamiento habitual es la derivación ventrículo-peritoneal, que conduce el exceso de LCR desde los ventrículos hacia la cavidad peritoneal.
