13.1. Desarrollo de las vellosidades coriónicas
El desarrollo de las vellosidades coriónicas es el proceso central de la placentación. Comienza en la segunda semana y alcanza su complejidad final hacia la semana 12. Se distinguen tres estadios.
13.1.1. Vellosidades primarias
Hacia el día 13-14, el citotrofoblasto prolifera y envía proyecciones hacia el interior del sincitiotrofoblasto, formando las vellosidades coriónicas primarias. Son estructuras simples: un núcleo de citotrofoblasto recubierto de sincitiotrofoblasto. No tienen vasos y el intercambio es únicamente por difusión a través de las capas de trofoblasto.
13.1.2. Vellosidades secundarias
Durante la tercera semana, el mesodermo extraembrionario crece hacia el interior del núcleo citotrofoblástico, transformando las vellosidades primarias en vellosidades secundarias. Ahora tienen un núcleo de mesodermo recubierto de citotrofoblasto y sincitiotrofoblasto. Todavía no tienen vasos fetales, pero el mesodermo central es el precursor del estroma vascular.
13.1.3. Vellosidades terciarias
A finales de la tercera semana, células angiogénicas del mesodermo central de las vellosidades secundarias se diferencian y forman capilares: son las vellosidades terciarias o vellosidades coriónicas maduras. Tienen un núcleo vascular (capilares fetales rodeados de estroma mesenquimal) recubierto de citotrofoblasto y sincitiotrofoblasto. Con la formación de las vellosidades terciarias se establece la circulación uteroplacentaria funcional: la sangre fetal circula por los capilares de las vellosidades, que están bañadas por sangre materna en los espacios intervellosos.
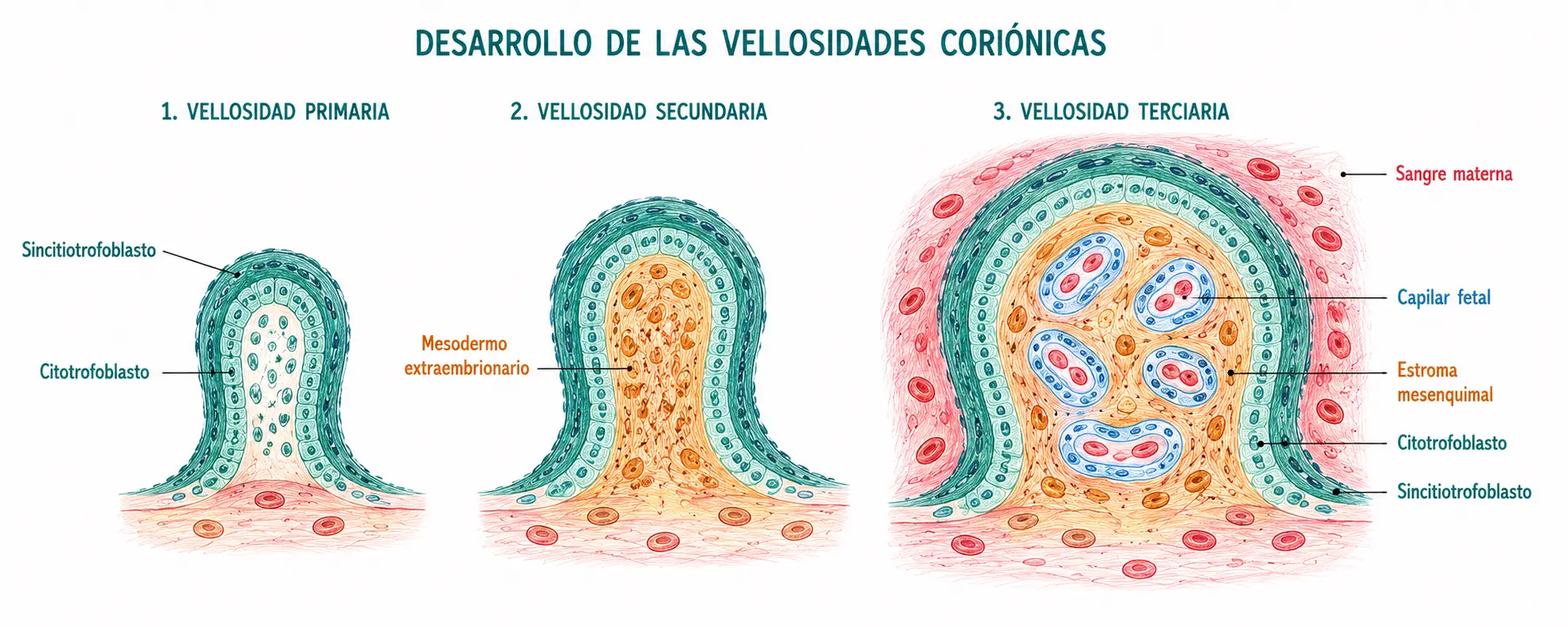
La secuencia primaria → secundaria → terciaria refleja tres eventos sucesivos:
- La invasión trofoblástica (primaria).
- La colonización mesenquimal (secundaria).
- La vascularización (terciaria).
Solo con la vascularización las vellosidades pueden realizar intercambio gaseoso real entre la madre y el feto. Antes de las vellosidades terciarias, la nutrición del embrión es por difusión simple y osmosis desde las secreciones glandulares endometriales (histiotrofia).
13.2. Estructura de la placenta madura
La placenta madura tiene forma de disco, mide unos 20 cm de diámetro y 3 cm de grosor, y pesa aproximadamente 500 g al término. Tiene dos componentes estructurales con dos orígenes distintos.
13.2.1. Componente fetal: la placa coriónica
La placa coriónica es la cara fetal de la placenta. Está formada por el corion frondoso: la región del corion (trofoblasto + mesodermo extraembrionario) que conserva sus vellosidades porque está en contacto con la decidua basal, la región más vascularizada del endometrio. El corion restante (que rodea la cavidad coriónica en las demás regiones) pierde sus vellosidades y forma el corion liso.
De la placa coriónica parten las vellosidades troncales (de anclaje), que se fijan a la decidua basal y de las que emergen las vellosidades libres (flotantes). Las vellosidades libres son las funcionalmente más activas: tienen una enorme superficie de intercambio y están bañadas por sangre materna.
13.2.2. Componente materno: la decidua basal
La decidua basal es la porción del endometrio situado entre el embrión y el miometrio. Forma el componente materno de la placenta. Está organizada en cotiledones. Los cotiledones son tabiques de tejido decidual que dividen el espacio intervelloso en compartimentos. Cada cotiledón contiene un conjunto de vellosidades troncales y sus ramificaciones.
Los espacios intervellosos son los espacios entre las vellosidades, dentro de cada cotiledón. Están llenos de sangre materna que entra directamente desde las arterias espirales del endometrio (erosionadas por el sincitiotrofoblasto) y sale por las venas endometriales. En estos espacios tiene lugar el intercambio materno-fetal.
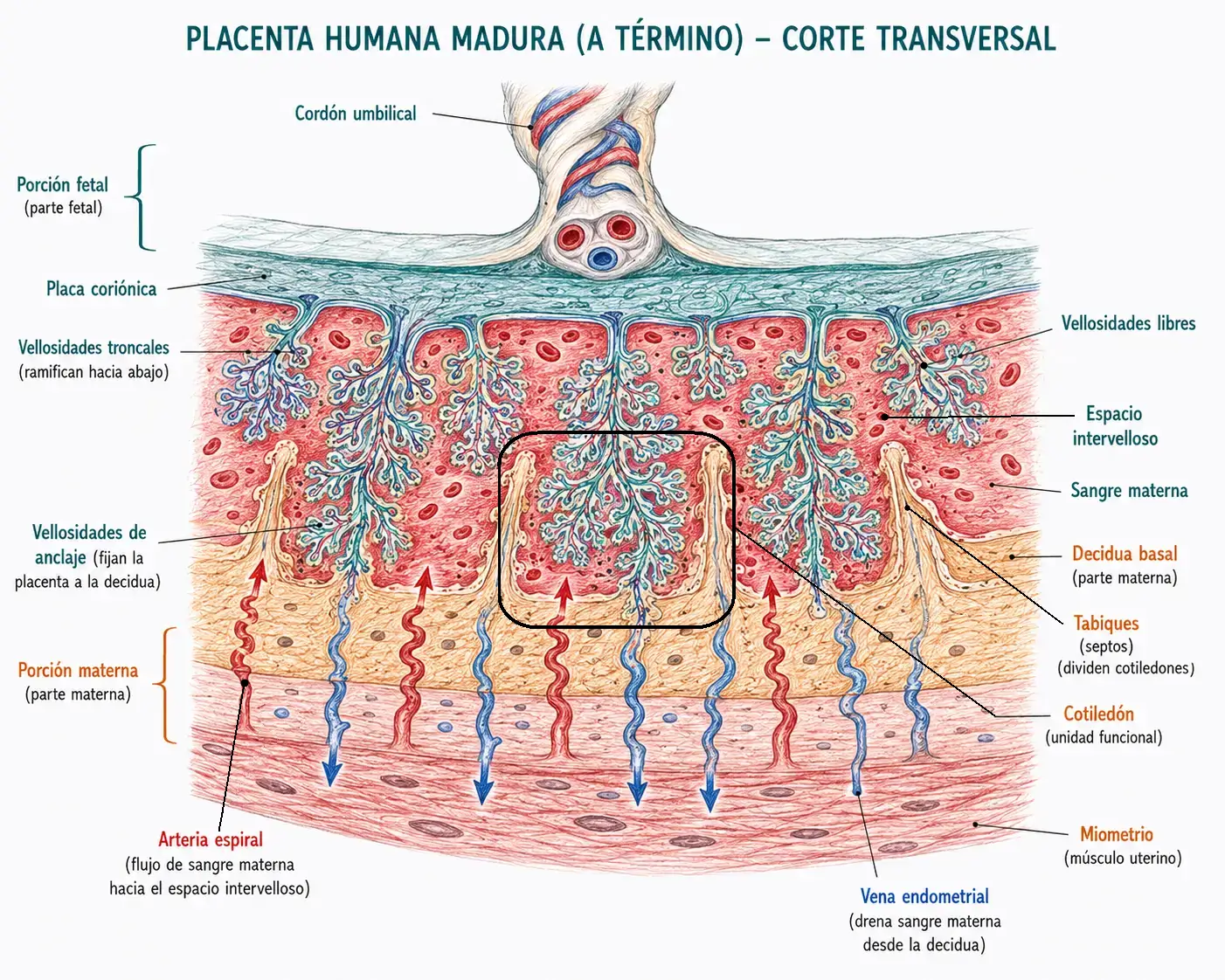
La sangre materna en los espacios intervellosos no circula a presión arterial completa: el sincitiotrofoblasto remodela las arterias espirales maternas convirtiéndolas en vasos de baja resistencia y baja presión. Si esta remodelación falla, la presión en los espacios intervellosos es excesiva y el flujo sanguíneo fetal se deteriora, contribuyendo a la preeclampsia y al retraso del crecimiento intrauterino.
13.2.3. La barrera placentaria
La barrera placentaria es el conjunto de capas que separan la sangre materna (en los espacios intervellosos) de la sangre fetal (en los capilares de las vellosidades). En el primer trimestre tiene cuatro capas: sincitiotrofoblasto, citotrofoblasto, estroma vellositario y endotelio capilar fetal. A medida que avanza el embarazo, el citotrofoblasto se hace discontinuo y el estroma se adelgaza, reduciendo el grosor de la barrera hasta 2-4 µm en el tercer trimestre.
La barrera placentaria no es impermeable. Permite el paso de oxígeno, glucosa, aminoácidos, ácidos grasos, vitaminas, agua e IgG maternas (inmunidad pasiva). También permiten pasar alcohol, nicotina, muchos fármacos, el virus de la rubéola, el citomegalovirus, el Toxoplasma gondii y el Treponema pallidum. El concepto erróneo de que la placenta "protege" al feto de todas las sustancias nocivas ha tenido consecuencias trágicas históricas (talidomida, síndrome de alcohol fetal).
13.3. Funciones de la placenta
La placenta tiene tres grandes categorías funcionales.
13.3.1. Intercambio materno-fetal
- El oxígeno y el CO₂ pasan por difusión simple favorecida por la alta afinidad de la hemoglobina fetal (HbF) por el oxígeno.
- La glucosa pasa por difusión facilitada.
- Los aminoácidos pasan por transporte activo.
- Las IgG maternas pasan por transcitosis mediada por receptor FcRn, dotando al feto de inmunidad pasiva frente a los patógenos a los que la madre ha sido expuesta.
- Los productos de desecho fetales (CO₂, urea, bilirrubina) pasan a la circulación materna para ser eliminados.
13.3.2. Función endocrina
La placenta es un órgano endocrino mayor que produce hormonas esenciales para el mantenimiento del embarazo.
- La hCG (gonadotropina coriónica humana) es la primera hormona placentaria, producida desde la implantación por el sincitiotrofoblasto. Mantiene activo el cuerpo lúteo durante las primeras 8-10 semanas, evitando la menstruación. Sus niveles alcanzan el máximo hacia la semana 10 y luego descienden. Es la base de los tests de embarazo.
- La progesterona es producida inicialmente por el cuerpo lúteo y, a partir de la semana 8-10, asumida por la placenta. Mantiene la quiescencia del miometrio, evita las contracciones prematuras y prepara el endometrio para la implantación.
- Los estrógenos placentarios (principalmente estriol) se producen mediante una cooperación entre la placenta y las suprarrenales fetales: la placenta carece de la enzima 16α-hidroxilasa, por lo que necesita precursores androgénicos (DHEA-S) procedentes de las suprarrenales fetales para sintetizar estriol. El estriol sérico materno es por tanto un indicador indirecto del bienestar de las suprarrenales fetales.
- El lactógeno placentario humano (hPL o somatomamotropina coriónica) modifica el metabolismo materno para asegurar el aporte de glucosa al feto: induce resistencia a la insulina materna (lo que eleva la glucemia materna disponible para el feto) y moviliza ácidos grasos maternos como fuente energética alternativa para la madre.
La diabetes gestacional es en parte consecuencia de la acción diabetogénica del hPL. En mujeres con reserva pancreática reducida, el hPL puede superar la capacidad compensatoria de las células beta, produciendo hiperglucemia materna. La hiperglucemia fetal que resulta estimula el páncreas fetal y produce hiperinsulinismo fetal, que a su vez causa macrosomía (feto grande), hipoglucemia neonatal al cortar el cordón y riesgo de complicaciones en el parto.
13.3.3. Función inmune
La placenta regula activamente la tolerancia inmunológica materna al feto. El sincitiotrofoblasto no expresa moléculas HLA clásicas (HLA-A, HLA-B) que serían reconocidas por los linfocitos T maternos. En su lugar expresa HLA-G y HLA-E, moléculas no clásicas que inhiben la actividad de las células NK maternas y promueven la tolerancia.
13.4. Las membranas fetales
Las membranas fetales son estructuras extraembrionarias que rodean y protegen al embrión o contribuyen a su nutrición. Son cuatro:
- Amnio.
- Corion.
- Saco vitelino.
- Alantoides.
13.4.1. El amnios
El amnios es la membrana más interna. Está formada por el epitelio amniótico (derivado del epiblasto) y una capa de mesodermo extraembrionario. Rodea completamente al feto y delimita la cavidad amniótica, que contiene el líquido amniótico.
El líquido amniótico se produce inicialmente por trasudación desde los vasos maternos y fetales. A partir del segundo trimestre, el feto deglute líquido amniótico (hasta 400-500 mL/día al término) y lo reabsorbe a través del intestino, y orina en la cavidad amniótica. Este ciclo continuo regula el volumen del líquido, que al término es de aproximadamente 800-1.000 mL.
El líquido amniótico tiene múltiples funciones:
- Protección mecánica contra traumatismos.
- Termorregulación.
- Permitir los movimientos fetales necesarios para el desarrollo musculoesquelético.
- Proporcionar un medio para el desarrollo pulmonar (el feto «respira» líquido amniótico, lo que expande los pulmones y es necesario para su maduración).
Las alteraciones del volumen de líquido amniótico tienen significado diagnóstico. El oligohidramnios (volumen reducido, ILA < 5 cm) puede indicar uropatía obstructiva o agenesia renal fetal (el riñón fetal produce la mayor parte del líquido en el segundo y tercer trimestre), rotura prematura de membranas o insuficiencia placentaria. El polihidramnios (volumen aumentado, ILA > 25 cm) puede indicar anomalías gastrointestinales fetales que impiden la deglución (atresia esofágica, anencefalia) o diabetes materna mal controlada.
13.4.2. El corion
El corion es la membrana más externa, formada por el trofoblasto y el mesodermo extraembrionario somatopleural. Rodea al embrión y al amnios. La región del corion en contacto con la decidua basal (corion frondoso) desarrolla vellosidades y forma la placa fetal de la placenta. La región opuesta (corion leve o liso) pierde sus vellosidades y queda como una membrana delgada avascular.
La fusión del amnios y el corion liso, que ocurre hacia la semana 14-16, forma las membranas amnio-coriónicas, que son las que se «rompen aguas» durante el parto.
13.4.3. El saco vitelino
El saco vitelino es una membrana derivada del hipoblasto (revestimiento) y del mesodermo extraembrionario esplacnopleural. En humanos no contiene vitelo nutritivo como en otros vertebrados, pero tiene funciones importantes durante el desarrollo temprano:
- Hematopoyesis: producción de células sanguíneas primitivas, a partir de la semana 3-4.
- Alberga inicialmente las células germinales primordiales que migrarán a las gónadas.
- Participa en la nutrición del embrión temprano mediante histiotrofia.
A medida que el embrión se pliega, el saco vitelino queda incorporado parcialmente al interior del embrión como el intestino primitivo. La conexión entre el saco vitelino externo y el intestino se estrecha formando el conducto vitelino (u onfalomesentérico), que normalmente se oblitera entre las semanas 5 y 9.
El divertículo de Meckel es el remanente más frecuente del conducto vitelino no obliterado. Presente en el 2% de la población, es un divertículo verdadero del íleon terminal situado a unos 60 cm de la válvula ileocecal. Puede contener mucosa gástrica ectópica (que produce ácido y causa úlcera y hemorragia) o mucosa pancreática ectópica. Se recuerda con la regla de los dos: 2% de prevalencia, 2 tipos de mucosa ectópica, 2 pies de distancia de la válvula ileocecal, 2:1 predominio en varones, sintomático en los primeros 2 años de vida.
13.4.4. La alantoides
La alantoides es una evaginación del saco vitelino hacia el pedículo de fijación que aparece hacia el día 16. En reptiles y aves tiene función de almacenamiento de desechos nitrogenados; en humanos es vestigial como estructura pero tiene importancia vascular: los vasos alantoideos forman las arterias y venas umbilicales que recorren el cordón umbilical y conectan la circulación fetal con la placenta.
La porción proximal de la alantoides persiste en el adulto como el uraco, que conecta el vértice de la vejiga urinaria con el ombligo. Normalmente se oblitera formando el ligamento umbilical medio.
13.5. El cordón umbilical
El cordón umbilical se forma a partir del pedículo de fijación (que contiene los vasos alantoideos y el conducto vitelino) recubierto por el amnios. Al término mide unos 50-60 cm de longitud y 1,5-2 cm de diámetro.
Contiene dos arterias umbilicales y una vena umbilical, embebidas en la gelatina de Wharton, un tejido conectivo laxo rico en ácido hialurónico que protege los vasos de la compresión. Las arterias umbilicales llevan sangre desoxigenada desde el feto hacia la placenta (a diferencia de las arterias sistémicas, que llevan sangre oxigenada: es el único ejemplo de arterias que llevan sangre venosa). La vena umbilical lleva sangre oxigenada desde la placenta hacia el feto.
La regla nemotécnica clásica para el contenido del cordón umbilical es AVA: dos Arterias y una Vena. También se recuerda que las Arterias llevan sangre Alejar (del feto hacia la placenta) y la Vena lleva sangre Volver (de la placenta al feto), aunque esto es contraintuitivo respecto a la nomenclatura vascular convencional.
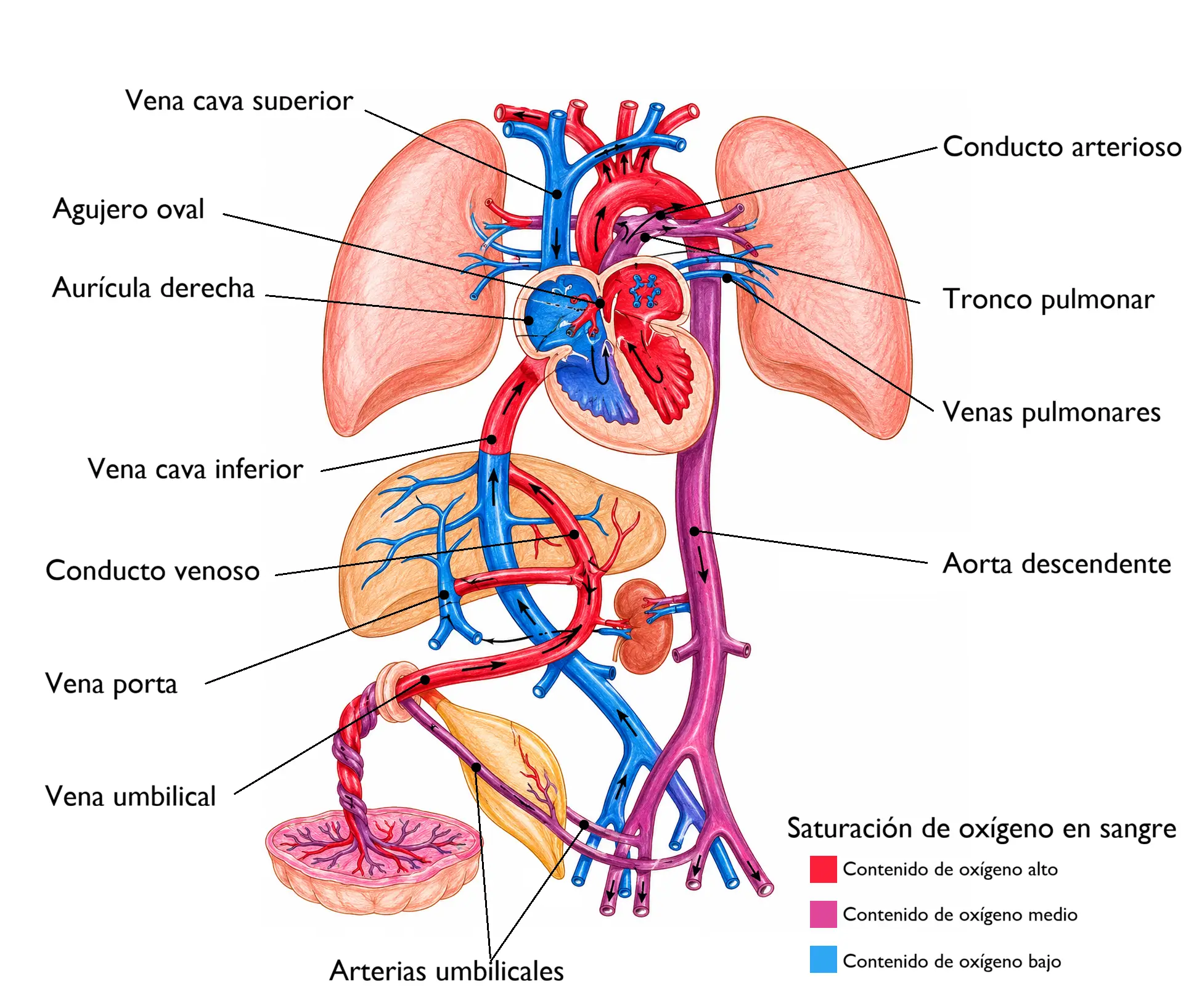
13.6. Circulación fetal
La circulación fetal es un circuito especializado que difiere fundamentalmente de la circulación adulta en tres aspectos:
- La placenta (no los pulmones) es el órgano de oxigenación.
- Los pulmones fetales no están expandidos y tienen alta resistencia vascular.
- Existen tres cortocircuitos que derivan la sangre oxigenada hacia los órganos prioritarios.
13.6.1. Recorrido de la sangre fetal
La sangre oxigenada procedente de la placenta llega al feto a través de la vena umbilical, que entra por el ombligo y se dirige al hígado. Una parte perfunde el hígado fetal a través de los sinusoides hepáticos. La mayor parte, sin embargo, se desvía directamente hacia la vena cava inferior a través del conducto venoso de Arancio, el primer cortocircuito fetal. Esto permite que la sangre más oxigenada alcance el corazón sin pasar por el hígado.
En la vena cava inferior, la sangre procedente del conducto venoso de Arancio se mezcla parcialmente con sangre desoxigenada procedente de las extremidades inferiores y el abdomen, aunque la crista dividens (un repliegue del tabique interauricular) la separa parcialmente al llegar a la aurícula derecha.
Desde la aurícula derecha, la sangre relativamente oxigenada es dirigida por la crista dividens a través del foramen oval hacia la aurícula izquierda, el segundo cortocircuito fetal. Desde la aurícula izquierda pasa al ventrículo izquierdo y de ahí a la aorta ascendente, irrigando con sangre bien oxigenada el corazón (arterias coronarias) y el encéfalo (arterias carótidas).
La sangre que permanece en la aurícula derecha (procedente de la vena cava superior, es decir, de la cabeza y los miembros superiores, ya desoxigenada) pasa al ventrículo derecho y de ahí a la arteria pulmonar. Sin embargo, debido a la alta resistencia vascular pulmonar de los pulmones no expandidos, la mayor parte de esta sangre no entra en los pulmones: es desviada hacia la aorta descendente a través del conducto arterioso (de Botal), el tercer cortocircuito fetal. Solo una pequeña fracción perfunde los pulmones.
Desde la aorta descendente, la sangre circula hacia las vísceras abdominales, las extremidades inferiores y, principalmente, hacia las arterias umbilicales, que la conducen de vuelta a la placenta para su reoxigenación.
| Cortocircuito | Conexión | Función | Equivalente adulto |
|---|---|---|---|
| Conducto venoso de Arancio | Vena umbilical → vena cava inferior | Deriva sangre oxigenada al corazón sin pasar por hígado | Ligamento venoso del hígado |
| Foramen oval | Aurícula derecha → aurícula izquierda | Deriva sangre oxigenada al ventrículo izquierdo y aorta | Fosa oval (tabique interauricular) |
| Conducto arterioso (de Botal) | Arteria pulmonar → aorta descendente | Deriva sangre del circuito pulmonar al sistémico | Ligamento arterioso |
13.6.2. Cambios circulatorios al nacimiento
En el momento del nacimiento se producen cambios fisiológicos rápidos que transforman la circulación fetal en circulación adulta. El desencadenante es la primera respiración.
La expansión pulmonar con la primera respiración produce una caída brusca de la resistencia vascular pulmonar. Esto aumenta el flujo sanguíneo hacia los pulmones y eleva la presión en la aurícula izquierda. Simultáneamente, el pinzamiento del cordón umbilical elimina el retorno venoso de la placenta, reduciendo la presión en la aurícula derecha.
El resultado es que la presión en la aurícula izquierda supera a la de la aurícula derecha: el foramen oval se cierra funcionalmente al quedar la válvula del foramen presionada contra el tabique. El cierre anatómico definitivo por fusión ocurre en los primeros meses de vida.
El aumento de la PO₂ arterial con la respiración desencadena la contracción del músculo liso del conducto arterioso, que se cierra funcionalmente en las primeras horas de vida. Su cierre anatómico definitivo (por proliferación de la íntima) ocurre hacia las 2-3 semanas. En el adulto persiste como el ligamento arterioso.
El conducto venoso de Arancio se cierra pasivamente al cesar el flujo de la vena umbilical tras el pinzamiento del cordón. En el adulto persiste como el ligamento venoso del hígado.
Las arterias umbilicales se contraen y se obliteran, persistiendo como los ligamentos umbilicales medios (porciones distales) y las arterias vesicales superiores (porciones proximales, que permanecen permeables irrigando la vejiga). La vena umbilical se oblitera formando el ligamento redondo del hígado (ligamento teres hepatis).
El ductus arterioso persistente (DAP) es el fallo en el cierre del conducto arterioso, más frecuente en prematuros (en los que los niveles de PO₂ son menores y la musculatura del conducto es inmadura). Produce un cortocircuito izquierda-derecha (de aorta a arteria pulmonar) que sobrecarga el circuito pulmonar. Se trata con indometacina o ibuprofeno (inhibidores de la COX que reducen las prostaglandinas vasodilatadoras que mantienen el conducto abierto) o con cierre quirúrgico o por cateterismo. La indometacina materna prenatal puede cerrar prematuramente el conducto in utero, razón por la que los AINEs están contraindicados en el tercer trimestre.
El foramen oval permeable (FOP) persiste en aproximadamente el 25% de la población adulta como un cierre incompleto del tabique interauricular. Habitualmente es asintomático, pero en ciertas situaciones (maniobra de Valsalva, embolismo paradójico) puede permitir el paso de trombos venosos a la circulación arterial, causando accidente cerebrovascular criptogénico. Es la causa más frecuente de ictus en adultos jóvenes sin factores de riesgo cardiovascular convencionales.
