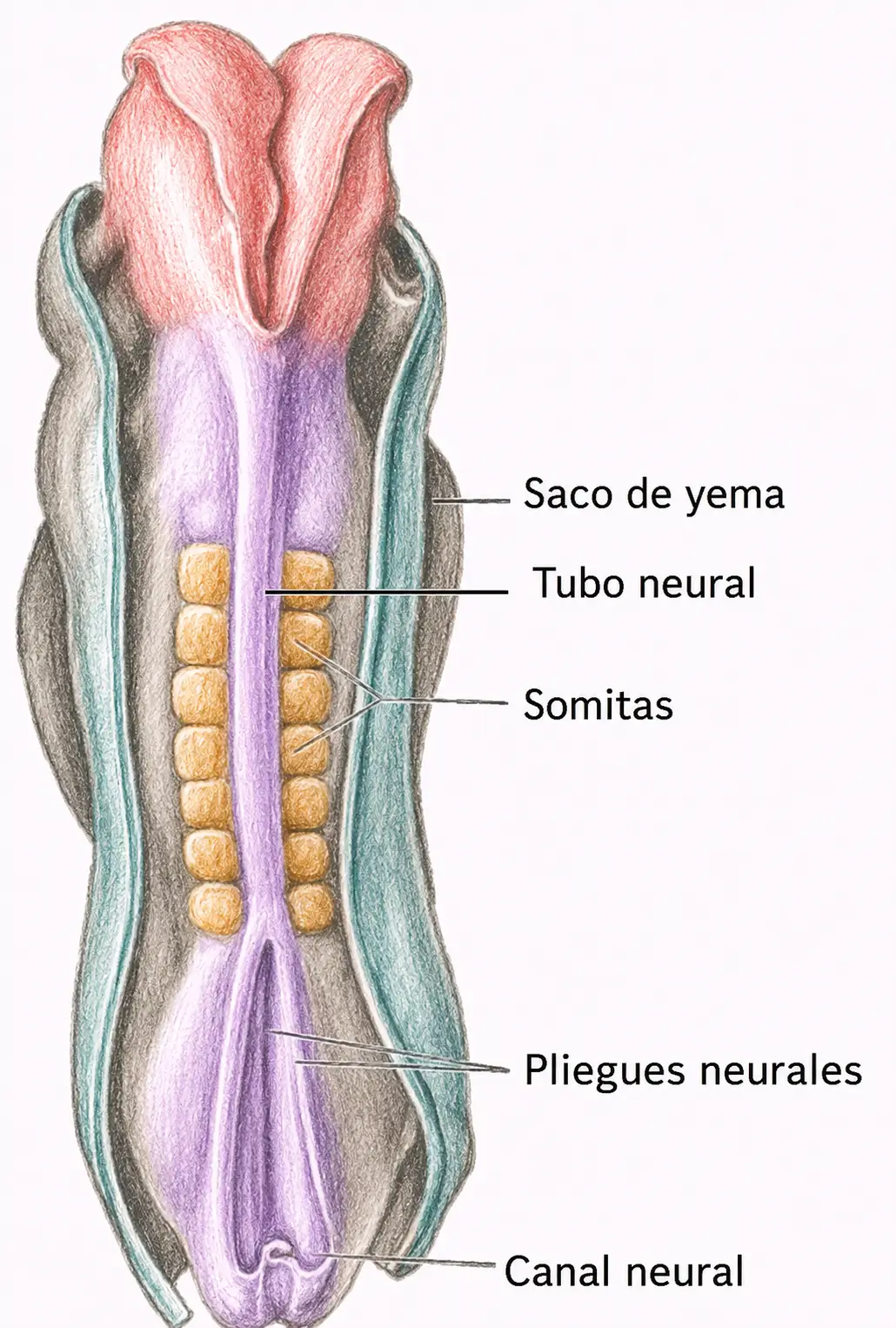
25.1. Introducción
Al final de la fase de gástrula quedan constituidos los somitas junto con las láminas que los recubren:
- La esplacnopleura por dentro.
- La somatopleura por fuera.
- La lámina intermedia (origen del aparato genital) entre ambas.
El embrión humano desarrolla entre 42 y 44 pares de somitas, distribuidos en:
- 4 occipitales: el primer somita occipital regresa antes de completarse la diferenciación somítica.
- 8 cervicales.
- 12 torácicos.
- 5 lumbares.
- 5 sacros.
- 8-10 coccígeos: los últimos 5-7 también regresan.
Debido a la regresión del somita occipital y los coccígeos, el número de metámeros funcionales en el adulto es menor que el número inicial de somitas formados.
La diferenciación de cada somita avanza en dirección cefalocaudal, no simultáneamente. En un mismo momento del desarrollo coexisten somitas ya diferenciados en la región cervical con otros todavía indiferenciados en la región lumbosacra.
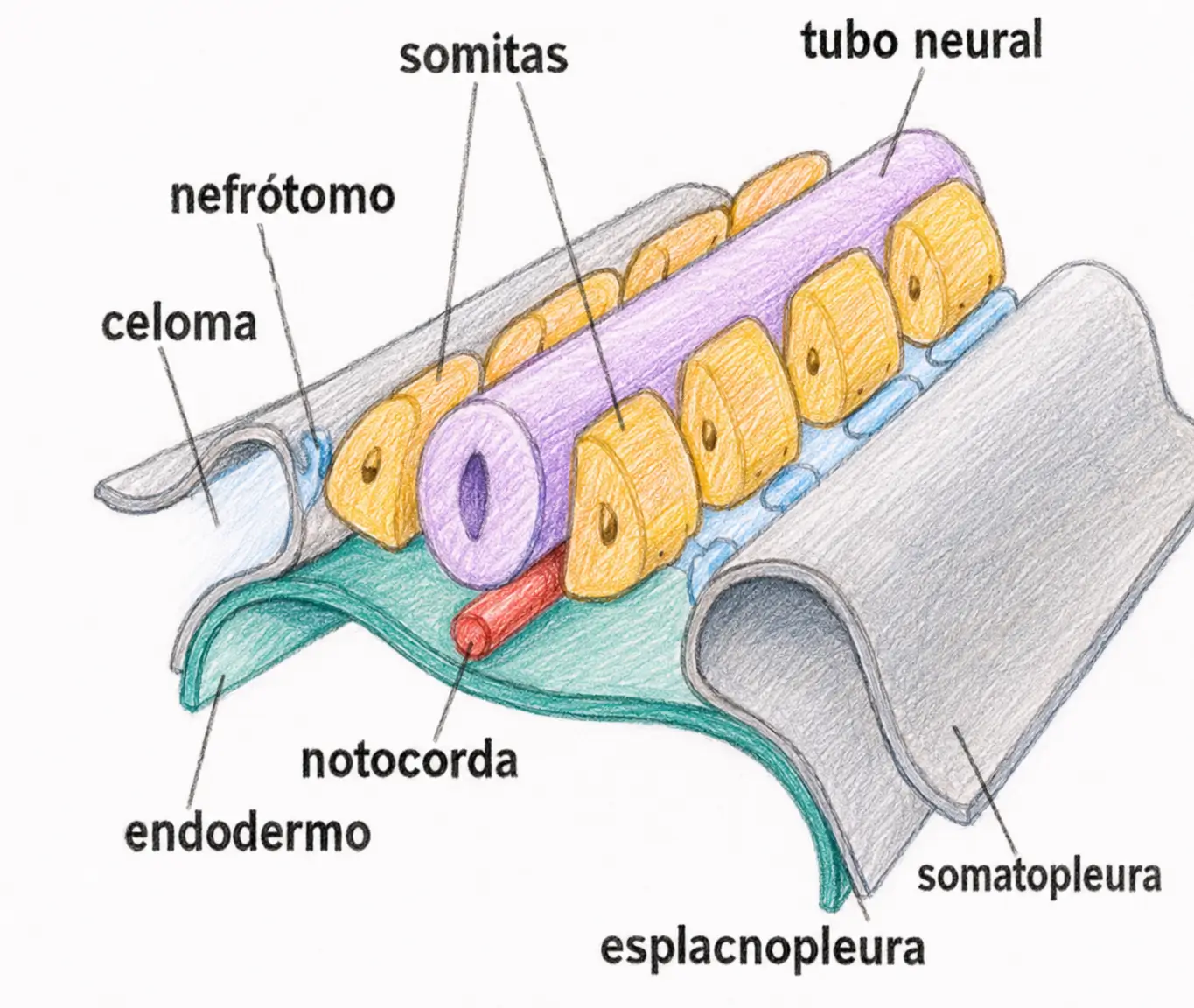
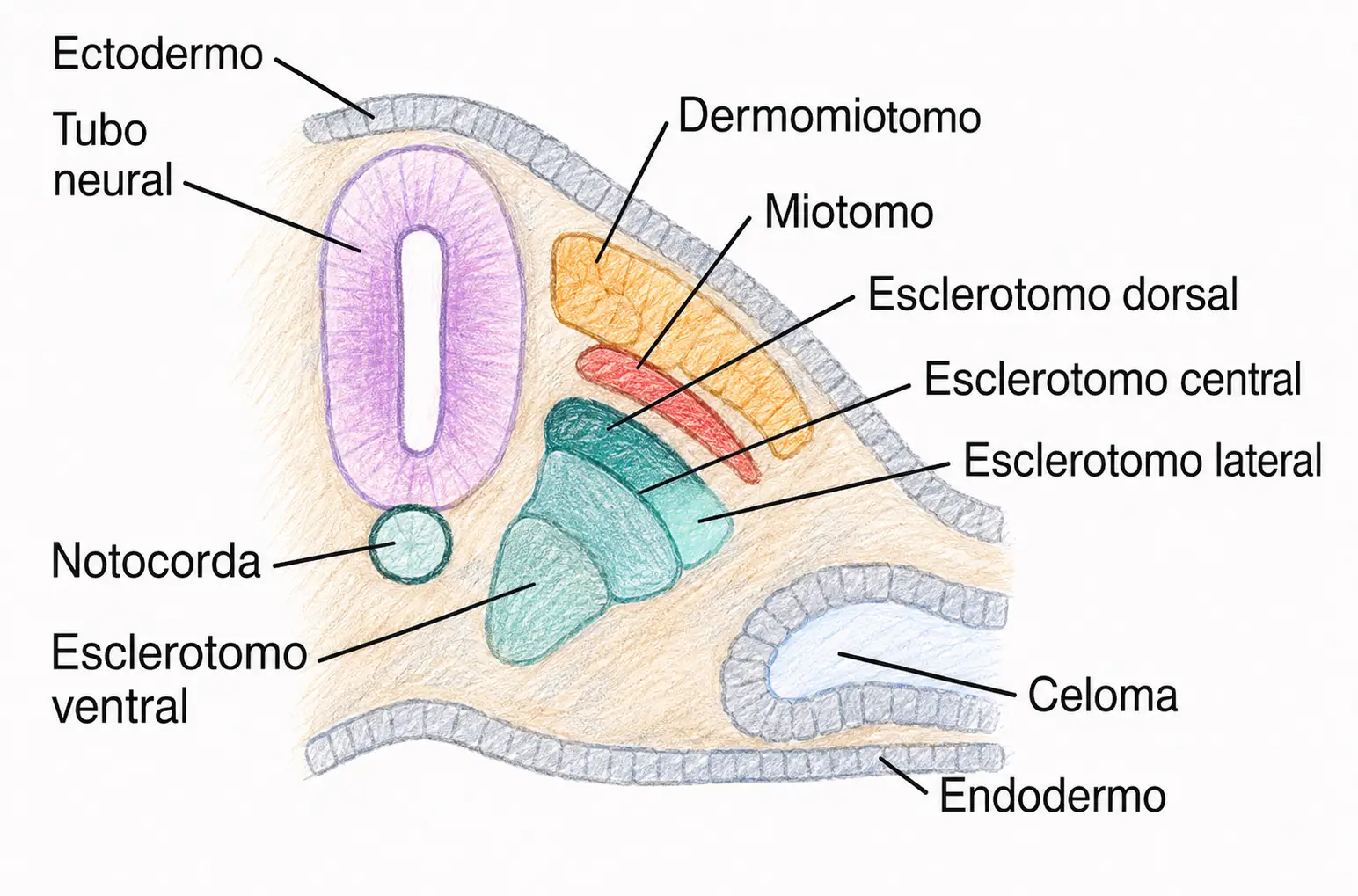
25.2. El somita general: esclerotomo, miotomo y dermotomo
25.2.1. Esclerotomo y resegmentación vertebral
La porción ventromedial del somita, el esclerotomo, es la responsable de formar el esqueleto axial. Sus células comienzan por depositar sales cálcicas (apatitas) antes de migrar en dos direcciones distintas:
- Una parte rodea la notocorda y el tubo neural formando el arco neural, que constituirá el cuerpo vertebral y el arco posterior de la vértebra.
- Otra parte migra hacia delante, sobre todo a nivel torácico, formando el arco visceral, origen de las costillas.
Cada esclerotomo se divide en una mitad craneal, laxa, y una mitad caudal, densa. La vértebra definitiva se forma a partir de la unión entre la mitad caudal de un esclerotomo y la mitad craneal del esclerotomo inmediatamente inferior. Este fenómeno se llama resegmentación.
La resegmentación tiene una consecuencia funcional directa. Al quedar la vértebra «desplazada» respecto al somita original, los miotomos correspondientes (y los nervios espinales que los inervan, que sí mantienen su nivel metamérico original) terminan situados a caballo entre dos vértebras consecutivas.
Esto es lo que permite que los músculos intervertebrales (por ejemplo, los intercostales) conecten dos vértebras adyacentes y no queden confinados dentro de una única vértebra.
25.2.2. Miotomo
La porción ventrolateral del somita, el miotomo, da lugar a fibras musculares con capacidad contráctil, que migran después hacia sus territorios definitivos para formar la musculatura del tronco y de las extremidades.
Según su comportamiento funcional, esta musculatura de origen somítico se clasifica en tres tipos:
- Lisa (involuntaria).
- Estriada o esquelética (mayoritariamente voluntaria).
- Miocárdica.
25.2.3. Dermotomo
La porción dorsal del somita, el dermotomo, se dispone bajo el ectodermo que formará la epidermis y constituye la dermis y la hipodermis (esta última vascularizada). De la piel derivan también sus anexos:
- Pelos.
- Uñas.
- Glándulas sebáceas y sudoríparas.
El ectodermo suprayacente, que forma la epidermis propiamente dicha (no el dermotomo), se estudia con detalle histológico en Histología Humana.
25.3. Formación de las extremidades
25.3.1. Niveles metaméricos y cresta ectodérmica apical
Las extremidades se originan a partir de metámeros específicos y fijos, cuya localización explica directamente la distribución de dermatomas y miotomos que persiste en el adulto:
- Los miembros superiores se forman desde el metámero C5 hasta D1.
- Los miembros inferiores, desde L2 hasta S3.
Cresta ectodérmica apical, o AER (apical ectodermal ridge): engrosamiento ectodérmico que aparece en el vértice del brote de la extremidad y actúa como centro señalizador del crecimiento en sentido proximodistal.
25.3.2. Control molecular de los tres ejes de la extremidad
El crecimiento y la organización espacial de la extremidad dependen de tres centros señalizadores independientes, cada uno responsable de un eje espacial distinto:
- El eje proximodistal (qué tan larga es la extremidad) depende de la AER y de la señalización FGF8/FGF10: mientras la AER se mantiene activa, el mesénquima subyacente sigue proliferando y alargando el miembro.
- El eje anteroposterior (qué dedo es cuál, pulgar frente a meñique) depende de la zona de actividad polarizante (ZPA), un grupo de células en el borde posterior del brote que secreta SHH (sonic hedgehog).
- El eje dorsoventral (dorso de la mano frente a palma) depende de la expresión de WNT7a en el ectodermo dorsal.
| Centro señalizador | Eje que controla | Molécula clave | Consecuencia del fallo |
|---|---|---|---|
| AER | Proximodistal (longitud) | FGF8 / FGF10 | Focomelia o amelia según el momento de pérdida de la señal |
| ZPA | Anteroposterior (identidad digital) | SHH (sonic hedgehog) | Duplicaciones especulares, polidactilia en espejo |
| Ectodermo dorsal | Dorsoventral (dorso/palma) | WNT7a | Ventralización del dorso de la extremidad |
La pérdida temprana de la AER detiene el crecimiento proximodistal en el punto exacto en que se pierde la señal, produciendo defectos por reducción longitudinal (de la focomelia a la amelia según el momento). La alteración de la señalización SHH/ZPA (por ejemplo, si se implanta tejido de ZPA en el borde anterior de un brote experimental) produce duplicaciones especulares de estructuras distales, como la polidactilia en espejo.
25.3.3. Las cuatro partes de la extremidad
Toda extremidad, superior o inferior, se organiza en las mismas cuatro partes con lógica proximal-distal:
- Basípodo (punto de unión al tronco).
- Estilópodo (brazo o muslo).
- Zigópodo (antebrazo o pierna).
- Autópodo o multípodo (mano o pie).
25.4. Malformaciones de las extremidades
25.4.1. Teratógenos: el caso de la talidomida
La talidomida ejerce su efecto teratógeno inhibiendo la angiogénesis del brote de la extremidad, mediante la degradación de la proteína SALL4 (y PLZF) a través de la vía del cereblon. Su acción es específica del periodo crítico de la cuarta a la séptima semana de gestación (formación activa de la extremidad); administrada fuera de esa ventana, no produce el mismo espectro de malformación. Según el día exacto de exposición dentro de esa ventana, el resultado varía entre focomelia (falta de un segmento proximal con partes distales presentes) y amelia (ausencia completa del miembro).
25.4.2. Malformaciones posicionales y óseas
No todas las malformaciones de extremidades tienen origen teratógeno o vascular. Algunas son defectos de posición mecánica y otras, defectos genéticos del propio tejido óseo o cartilaginoso.
El pie en calzador (arco interno convexo) y el pie en astrágalo vertical (detectable hacia la semana 20) son defectos posicionales que dificultan la deambulación. Los defectos por plegamiento excesivo del miembro sin la extensión adecuada dan lugar a clinodactilia.
La acondroplasia es una condrodisplasia de origen genético, causada por mutación activadora del gen FGFR3, que produce un fallo de proliferación en el cartílago de crecimiento y, en consecuencia, talla baja con acortamiento rizomélico (predominantemente proximal) de las extremidades.
La osteogénesis imperfecta es un trastorno genético del colágeno tipo I que produce fragilidad ósea. Los huesos se fracturan con facilidad tanto en la vida intrauterina como extrauterina; es frecuente que el recién nacido presente ya callos de fractura formados por la simple compresión mecánica del útero.
El pie equino es diagnosticable antes del nacimiento pero no remediable en esa etapa; la resección del tendón de Aquiles puede ser beneficiosa en su manejo posnatal. No debe confundirse con el pie en calzador ni con el pie en astrágalo vertical, que son defectos de forma, no de tensión tendinosa.
